
Эффект Варбурга: какие преимущества он даёт опухолевым клеткам?

Метаболизм опухолевых клеток тесно связан с их долгосрочным ростом, способностью к выживанию, пролиферацией. Отличительная черта такого метаболизма заключается в повышенном потреблении глюкозы и ферментативном превращении глюкозы в лактат. Этот феномен имеет место при полностью функционирующих митохондриях и носит название эффект Варбурга. Существование этого эффекта предполагалось около 90 лет назад, но изучить его должным образом удалось лишь 10 лет назад — на тысячах страниц были описаны причины и функции данного феномена. Несмотря на высокий интерес, природа эффекта Варбурга остаётся неизвестной. В этой статье проанализированы некоторые представления о роли данного феномена с особым вниманием к обоснованию и обсуждением противоречий.
Основные моменты:
- И гликолитический, и митохондриальный метаболизм необходимы для реализации клеточной пролиферации — как в прошлом понимании эффекта Варбурга, так и в настоящем.
- За многие годы появилось немало предположений о предназначении эффекта Варбурга. Каждая из предполагаемых функций по-своему привлекательна, но также поднимает и определённые вопросы.
- Функции сигнальной передачи кажутся весьма вероятными относительно эффекта Варбурга, однако это трудно проверить экспериментально.
Глоссарий
- Аэробный гликолиз: усиленная интенсивность гликолиза и ферментативное превращение до лактата, происходящие при условии нормально функционирующих митохондрий.
- АТФ: аденозинтрифосфат, разменная монета энергетического обмена клетки.
- Поток: суммарная скорость химической реакции, результирующая из превращения одного метаболита в другой при его прохождении по определённому метаболическому пути.
- NADH: восстановленная форма никотинамидадениндинуклеотида (NAD+); восстановитель, принимающий участие в ОВР, лежащих в основе транспорта электронов. NADH является ключевым восстановительным эквивалентом в реакциях гликолиза, а также в реакциях, протекающих в митохондриях.
- NADPH: восстановленная форма никотинамидадениндинуклеотидфосфата (NADP+); привлекается для восстановительного биосинтеза и регенерации восстановленного глутатиона.
- Активные формы кислорода: восстановленные формы кислорода, обладающие химически агрессивными свойствами.
- Эффект Варбурга: альтернативное название аэробного гликолиза, введённое Эфраимом Рекером в начале 1970-х.
Метаболизм глюкозы и эффект Варбурга
Метаболизм одного из главных макронутриентов — глюкозы — позволяет вырабатывать энергию в виде АТФ посредством окисления и образования углеродных связей. Этот процесс является ключевым в жизни всех млекопитающих. Конечным продуктом может являться лактат или, при полном окислении глюкозы, CO2. В опухолях и других пролиферирующих клетках уровень поглощения глюкозы резко повышен, вследствие чего производится лактат даже в присутствии кислорода и функционирующей митохондрии. Этот процесс, названный эффектом Варбурга, был глубоко изучен.
Однако после более тщательного анализа стало очевидным то, что вопросы преимущества роста клеток и их способности к выживанию до сих пор не решены. В этом анализе было уделено особое внимание нескольким предположениям об их функциях и в каждом случае разобраны возникающие вопросы. Перед представлением различных аспектов эффекта Варбурга сначала обсудим исторический контекст с ударением на концептуальное развитие.
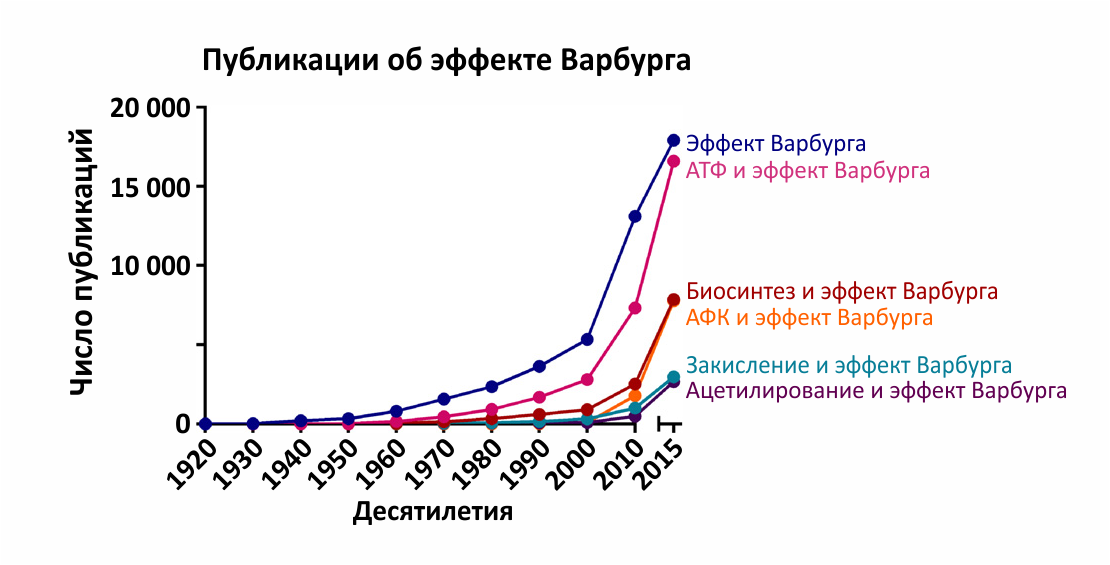
Рисунок 1 | Зависимость количества публикаций на тему эффекта Варбурга от времени публикации.
Исторические предпосылки к изучению эффекта Варбурга
В 1920-х годах Отто Варбург и его коллеги доказали в своём исследовании, что опухоли использовали необычайно большое количество глюкозы по сравнению с клетками здоровой окружающей ткани. Кроме того, глюкоза была подвержена нескольким ферментативным реакциям, в ходе которых образовался лактат даже в присутствии кислорода. Так произошёл термин аэробный гликолиз [1,2]. Однако, было отмечено, что клеточное дыхание само по себе может поддерживать жизнеспособность опухоли. Был сделан вывод, что для избавления от опухолевых клеток путём лишения их энергии было необходимо устранить подачу как глюкозы, так и кислорода [3]. Позже, в 1929 году, английский биохимик Герберт Крабтри дополнил работу Варбурга исследованием гетерогенности гликолиза в клетках опухоли. Он подтвердил выводы Варбурга, но также обнаружил, что интенсивность дыхания в опухолях была переменной, при этом многие опухоли имели значительно большую интенсивность дыхания [4]. Таким образом, Крабтри пришел к выводу, что опухолевые клетки не только обладают аэробным гликолизом, но существует также вариабельность в скорости ферментации, по-видимому, из-за экологических и органических влияний.
Вопреки результатам предыдущих работ и по причинам, неясным для авторов, Варбург позднее предположил, что нефункционирующие митохондрии являются причиной аэробного гликолиза [5]. Далее Варбург выдвинул гипотезу о том, что именно этот процесс и является причиной развития рака. В ранних 1970-х Эфраим Рекер предложил назвать феномен в честь его открывателя — “эффект Варбурга”. Рекер развил свою собственную теорию о происхождении Эффекта Варбурга, основанную на дисбалансе внутриклеточного pH, влияющем на активность АТФ-азы [6]. Джеффри Флир, Моррис Бёрнбаум и Эфраим Рекер обнаружили, что аэробный гликолиз является контролируемым процессом, который может непосредственно регулироваться фактором роста. К этому времени открытие онкогенов привело к выводу, что аберрантная регуляция передачи факторов роста является инициирующим событием в онкогенезе. Таким образом, их наблюдения принесли новое значение гипотезы Варбурга в биологии рака [7–10]. Тем не менее оставалась неясна причинно-следственная связь между развитием рака и эффектом Варбурга до тех пор, пока генетические и фармакологические исследования убедительно не показали, что эффект Варбурга первичен по отношению к росту опухоли [11,12]. Сейчас ясно, что опухолевый метаболизм затрагивает две составляющие: аэробный гликолиз и митохондриальный метаболизм [13–16]. На протяжении всей истории назначение эффекта Варбурга оставалось спорным. В этой статье будут представлены основные предположения на этот счёт.
Эффект Варбурга и быстрый синтез АТФ
При пересчёте на единицу глюкозы аэробный гликолиз представляется неэффективным способом получения АТФ, если сравнивать с количеством молекул АТФ, синтезируемых в ходе этапа клеточного дыхания, протекающего в митохондриях [17,18]. Тем не менее интенсивность метаболизма глюкозы по пути аэробного гликолиза выше, и таким образом, образование лактата из глюкозы осуществляется в 10‒100 раз быстрее, чем полное окисление глюкозы в митохондриях. Фактически, количество АТФ, синтезируемое в любой взятый промежуток времени, сопоставимо, какая бы из форм метаболизма глюкозы ни была использована [19]. Таким образом, разумная гипотеза о том, для чего клетки будут прибегать к аэробному гликолизу, объясняла бы имеющееся различие в кинетике.
Теоретические расчёты с использованием эволюционной теории игр подтверждают гипотезу о том, что клетки с более высокой интенсивностью гликолиза, но меньшим выходом АТФ могут получать эволюционное преимущество, когда имеет место конкуренция за долю ограниченных энергетических ресурсов (рис. 2) [20,21]. По сути, опухолевое микроокружение ограничивает доступность глюкозы и вступает в борьбу за питательные вещества со стромальными клетками и клетками иммунного звена [22,23].
Дополнительная информация по этому поводу была получена из недавнего исследования, показавшего, что в случае если сдвиги в клеточной среде приводили к существенному увеличению потребности в АТФ, обусловленному энергетическими изменениями АТФ-зависимых мембранных насосов, то наблюдалось усиление аэробного гликолиза, тогда как окислительное фосфорилирование оставалось на неизменном уровне [24]. Полученные данные являются ещё одним подтверждением в пользу того, что роль эффекта Варбурга заключается в поддержании скорейшего пути синтеза АТФ, который может быстро перестраиваться в зависимости от спроса на АТФ.
Несмотря на столь привлекательный план, имеется немало трудностей. Простые эмпирические расчёты указывают на то, что требуемое для роста и размножения клетки количество АТФ может быть меньше, чем в норме требуется для жизнедеятельности [18,25]. Так, энергетические запросы могут и вовсе не достигать критического значения во время роста опухолевой клетки. Более того, механизмы, доступные для других типов клеток в случае экстренной нужды в АТФ, также имеются в распоряжении и у опухолевых клеток. Например, быстрое получение АТФ за счёт креатинкиназ в работающих мышцах или аденилаткиназы под влиянием гормональных сдвигов происходит почти в большинстве опухолевых клеток и должно бы отвечать потребностям в АТФ. Потому необходимы дальнейшие исследования, чтобы продемонстрировать, действительно ли этот механизм лежит в основе аэробного гликолиза.
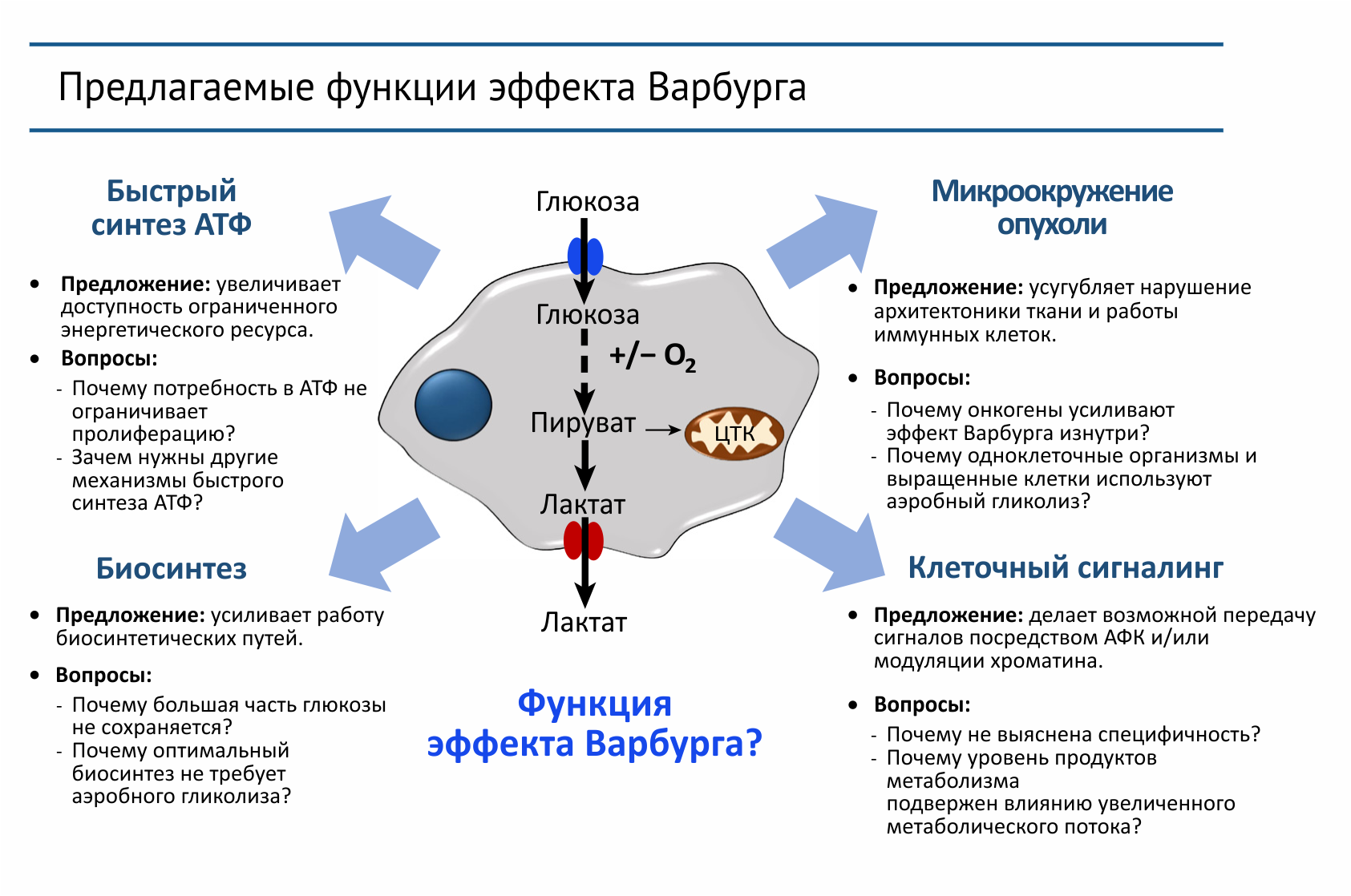
Рисунок 2 | Эффект Варбурга заключается в увеличении захвата глюкозы клеткой и преимущественной продукции лактата даже в присутствии кислорода. Каждая из этих функций была представлена как функция эффекта Варбурга.
Сокращения: АФК — активные формы кислорода, ЦТК — цикл трикарбоновых кислот.
Эффект Варбурга и биосинтез
Эффект Варбурга был предложен как адаптационный механизм для поддержки биосинтетических требований к неконтролируемой пролиферации (рис. 2). В этом сценарии повышенное потребление глюкозы используется в качестве источника углерода для анаболических процессов, необходимых для поддержки клеточной пролиферации [17,26–32]. Этот избыток углерода может участвовать в многочисленных ответвлениях гликолиза и используется для создания нуклеотидов, липидов и белков. Одним из примеров является использование гликолитического потока в биосинтезе серина de novo через фермент фосфоглицератдегидрогеназу (PHGDH) [18]. Вместо того, чтобы зависеть от ограничивающей скорость потребности в АТФ, пролиферирующие клетки больше нуждаются в восстановленных эквивалентах в виде NADPH в дополнение к использованию дополнительного углерода от изменённого метаболизма глюкозы для построения клеточных элементов. Увеличение поглощения глюкозы позволяет более эффективно синтезировать эти восстановленные эквиваленты в окислительной ветви пентозофосфатного пути, которые затем используются в редуктивном биосинтезе, наиболее заметным в синтезе липидов de novo [17,33].
Другим предложенным механизмом использования эффекта Варбурга в биосинтезе является регенерация NAD+ из NADH на стадии преобразования пирувата до лактата, которая завершает аэробный гликолиз. В этом случае NADH, который продуцируется глицеральдегидфосфатдегидрогеназой (GAPDH), должен быть израсходован для регенерации NAD+, чтобы поддерживать гликолиз. Высокая скорость гликолиза позволяет линиям питания оставаться открытыми, что может, например, перенаправлять 3-фосфоглицерат (3PG) в серин для одноуглеродного опосредованного метаболизмом производства NADPH и нуклеотидов [17,25]. Из всех этих предложений можно заключить, что эффект Варбурга поддерживает метаболические процессы, позволяющие ускоренному биосинтезу поддерживать рост и пролиферацию.
Кроме этого, существуют предположения, что аэробный гликолиз является компромиссом для поддержания биосинтеза [34,35,62]. В этих условиях неэффективный способ получения АТФ является платой за поддержание высоких уровней анаболических процессов. Эти процессы требуют повышенной экспрессии генов биосинтеза, вовлечённых в нуклеотидный и липидный обмен, и компромисс происходит за счёт ограничения использования митохондрий для сохранения высокой экспрессии биосинтетических ферментов в условиях количественной ограниченности белков, которые можно произвести. Другим сценарием подобного компромисса является идея того, что физический объём, доступный для клетки, может ограничивать число митохондрий, и таким образом, любые потребности в энергии и биомассе, которые превышают ограниченную митохондриальную ёмкость, необходимо получать из аэробного гликолиза [36–38]. Эта концепция получила название «ограничение платёжеспособности». В обоих этих случаях эффект Варбурга является адаптацией к поддержке производства биомассы в ограниченных для синтеза АТФ условиях.
Привлекательность этой идеи частично объясняется её способностью давать простое объяснение очевидной корреляции между аэробным гликолизом и клеточным ростом/пролиферацией. Более того, кажется интуитивным, что вторичные ответвления от гликолиза будут использоваться в большей степени во время эффекта Варбурга, поскольку скорость гликолиза выше, а производство лактата в этом случае будет служить для регенерации NAD+, необходимого для поддержки гликолиза. Также необходимость NADPH для генерации липидов можно суммировать в очень простом химическом уравнении, показывающем, что потребность в NADPH выше, чем потребность в АТФ для биосинтеза [17].
Однако существуют серьёзные ограничения для данной предложенной функции эффекта Варбурга. Во-первых, во время аэробного гликолиза большая часть углерода не сохраняется, а вместо этого выводится в виде лактата [25]. На самом деле, общее уравнение одной молекулы глюкозы, превращаемой в две молекулы лактата без общего увеличения или потери NAD+ и NADH, не оставляет места для биомассы. Это происходит из-за стехиометрии гликолиза, где производство биомассы является взаимоисключающим с производством лактата и регенерация NAD+ для биосинтеза только за счёт лактата не может быть объяснена. Таким образом, пути, которые приводят к биосинтезу из глюкозы, происходят в полном отсутствии лактата, что является отличительной чертой эффекта Варбурга. Кроме того, широко известно, что митохондрии являются ключевыми компонентами программ биосинтеза, субстраты которых забираются из цикла трикарбоновой кислоты (TCA) и используются для биосинтеза нуклеотидов, аминокислот и липидов [39,40]. В свете этих данных остаётся трудным понять, каким образом эффект Варбурга может непосредственно способствовать биосинтезу.
Что касается предложений, которые определяют эффект Варбурга как компромисс для усиления биосинтеза, то недавние оценки количественных протеомных исследований показывают, что стоимость производства белка для проведения аэробного гликолиза огромна. Фактически клетки выделяют до 10% от всего своего протеома и половину всех своих метаболических генов, чтобы продуцировать белки, участвующие в гликолизе [41]. Биосинтетические программы в клетках, наоборот, требуют меньшего количества белка. Таким образом, стоимость производства белков для аэробного гликолиза столь же велика, если не больше, чем стоимость производства белков для биосинтеза. Эти предложения дополнительно оспариваются доказательствами, показывающими, что митохондриальные функции проходят одновременно с эффектом Варбурга, и таким образом, ограничения активности митохондрий во время эффекта Варбурга, по-видимому, не происходит. В конечном счёте необходимы дальнейшие исследования, чтобы выяснить, способствует ли эффект Варбурга поддержанию биосинтетических программ.
Эффект Варбурга и микроокружение опухоли
Отдельно от внутренних функций клетки, описанных в предыдущих разделах, эффект Варбурга может представлять преимущество для роста клеток в многоклеточной среде. Закисление микроокружения и другие метаболические эффекты — довольно интригующие возможности. Повышенный метаболизм глюкозы снижает рН в микроокружении вследствие секреции лактата (рис. 2) [42].
Потенциальные преимущества ацидоза для раковых клеток многообразны. Гипотеза кислотноопосредованной инвазии предполагает, что ионы водорода H+, выделяемые раковыми клетками, диффундируют в окружающую среду и изменяют характеристики опухолевой стромы, что обеспечивает усиленную инвазивность [42,43]. Недавнее исследование показало, что лактат, выделяемый опухолями, является источником поляризации тканевых макрофагов М2-типа [44]. Кроме того, доступность глюкозы, по-видимому, является результатом прямой конкуренции между опухолью и лимфоцитами, инфильтрирующими опухоль (TIL) [22,23]. Высокие уровни гликолиза ограничивают доступность глюкозы к TIL, которые требуют большого количества глюкозы для своего функционирования. В поддержку этого предположения есть доказательство того, что направленное воздействие на аэробный гликолиз опухоли имеет дополнительную выгоду в виде увеличения доступности глюкозы для TIL, увеличивая их противоопухолевые функции.
Вместе эти данные свидетельствуют о том, что опухолевые клетки могут взаимодействовать с клетками иммунной системы для поддержания условий иммунитета, необходимых для опухоли.
Вероятно, эффект Варбурга поддерживает микроокружение опухоли, способствующее пролиферации раковых клеток. Тем не менее считается, что эффект Варбурга является ранним событием в онкогенезе как следствие начальной онкогенной мутации, такой как мутации KRAS при раке поджелудочной железы или мутации BRAF при меланоме; таким образом, он происходит не только до инвазии клеток, но и при доброкачественных и ранних стадиях поражения [45,46]. Другая проблема заключается в том, что в условиях, полностью изолированных от окружающей среды, таких как фаза роста одноклеточных дрожжей, эффект Варбурга остаётся выбором углеводного метаболизма [38]. Совместно эти данные указывают на то, что не-внутриклеточных функций эффекта Варбурга недостаточно для полного объяснения его функций.
Эффект Варбурга и клеточный сигналинг
Мы и другие исследователи предполагаем, что эффект Варбурга прямо влияет на сигнальные функции опухолевых клеток [18,39,47–49]. Это предположение особенно привлекательно благодаря определению непосредственной причинной роли изменённого метаболизма глюкозы в поддержке онкогенеза как результата изменённой трансдукции сигнала, оказывающей влияние на клеточные процессы. Двумя областями работы сигналинга являются генерация и регуляция уровня активных форм кислорода (АФК), а также регуляция состояния хроматина. Другие исследования выявили дополнительные возможные механизмы сигналинга [23,50]. Жизненно важным является поддержание баланса АФК [51]. Избыток АФК повреждает клеточные мембраны и нуклеиновые кислоты, а также оказывает другие разрушительные эффекты. Дефицит АФК нарушает процессы сигналинга, необходимые для клеточной пролиферации, например, путём инактивации гомолога фосфатазы и тензина (phosphatase and tensin homolog, PTEN) и тирозиновых фосфатаз.
Эффект Варбурга вызывает изменение редокс-потенциала митохондрий, необратимо приводящее к изменениям в генерации АФК [18]. Важным показателем редокс-потенциала в клетках является NADH, который участвует в переносе электронов в митохондриях. Клеточные механизмы, поддерживающие редокс-гомеостаз, включаются при резких изменениях интенсивности гликолиза. До определённого уровня интенсивности гликолиза малат-аспартатный шунт способен компенсировать дефицит NADH из митохондрий [18]. Однако, когда скорость гликолиза превышает возможности малат-аспартатного шунта, восстановление NAD+ становится возможным путём превращения пирувата в лактат при помощи лактатдегидрогеназы (ЛДГ). Этот процесс также может влиять на производство АФК путём изменения концентрации восстановленных эквивалентов в митохондриях (рис. 2) [18,52]. Это последствие эффекта Варбурга может быть прямо вовлечено в онкоген-индуцированное старение (ОИС) [53]. ОИС имеет онкосупрессивную клеточную функцию, и недавно стало известно, что повышенная интенсивность окисления глюкозы через пируватдегидрогеназу может оказывать влияние на ОИС. Это показало, что редокс-баланс NADH может быть вовлечён в прямые сигнальные роли эффекта Варбурга.
Кроме того, метаболические пути, стимулирующие редокс-гомеостаз, активируются (путём апрегуляции) вместе с эффектом Варбурга. Например, пентозофосфатный путь, происходящий из гликолиза, производит NADPH, а биосинтез серина de novo, снабжающий метаболизм одноуглеродных остатков, производит NADPH и глутатион, регулирующий уровень АФК [54,55]. Вместе эти открытия дают прямую биохимическую связь между аэробным окислением глюкозы и доступностью АФК, которые, в свою очередь, могут влиять на множество сигнальных путей.
Кроме клеточного сигналинга при помощи АФК, была обнаружена связь между метаболизмом глюкозы и ацетилированием гистонов [56–59]. Состояние хроматина отвечает за различные функции клеток, в том числе репарацию ДНК и транскрипцию генов. Было установлено, что уровень ацетил-КоА, субстрата для ацетилирования гистонов, регулируется притоком глюкозы [59]. Была показана прямая связь между клеточным метаболизмом и регуляцией генов роста, а также возможность внутриклеточных уровней ацетил-КоА быть высококонсервативным механизмом, который обеспечивает эту важную связь [60]. Активность АТФ-цитратлиазы, фермента, отвечающего за превращение цитрата в ацетил-КоА (и оксалоацетат, прим. перев.), может влиять на уровень ацетилирования гистонов [47]. Повышение уровня ацетил-КоА может быть достаточным для того, чтобы путём ацетилирования гистонов переключить клетки в фазу роста [56]. Удаление глюкозы или снижение активности АТФ-цитратлиазы снижает ацетилирование на ряде гистонов и уменьшает транскрипцию генов, вовлечённых в метаболизм глюкозы. Это значит, что есть некоторая взаимосвязь между метаболизмом глюкозы и ацетилированием гистонов. Согласно этой идее, метаболизм глюкозы оказывает влияние на структуру хроматина [58]. Кроме ацетилирования гистонов в ответ на доступность глюкозы, деацетилирование также может быть реакцией в ответ на доступность нутриентов [39]. Деацетилирование играет важную роль в чувствительности клеток к нутриентам и сигналинге, так как активность многих деацетилаз зависит от уровня NAD+. Отношение NAD+/NADH увеличивается при состояниях с недостатком нутриентов [39,56,57]. Поэтому ацетилирование и деацетилирование могут регулироваться доступностью нутриентов, и это говорит о том, что эти состояния могут быть следствием эффекта Варбурга. Эти доказательства свидетельствуют о сигнальных функциях гликолиза.
Однако, ряд трудностей ограничивает возможность предложенных функций быть главным механизмом, приносящим пользу раковым клеткам путём переключения на аэробный гликолиз. Во-первых, трудно представить, каким образом реализуется молекулярная специфичность в таком большом и обширном механизме передачи сигналов. В противоположность этому стоит, например, сигналинг факторов роста, в котором лиганд, связываясь с субстратом, вызывает изменения конформации и ферментативной активности, приводящие к изменениям определённых клеточных процессов. Однако механизму, посредством которого состояние гликолиза отдаёт сигнал другим клеточным процессам, не хватает очевидных субстратов специфичности.
Другим ограничением является то, что такие предложения обычно не обладают фальсифицируемостью. Это значит, что трудно придумать эксперимент, который бы окончательно показал, что специфический сигнальный механизм, такой как изменение структуры хроматина, является прямым результатом состояния метаболизма глюкозы как основного преимущества аэробного гликолиза.
Ещё одним затруднением является то, что биохимические реакции проходят быстро, а фенотипические клеточные изменения происходят на протяжении больших промежутков времени, что допускает дополнительное вмешательство многих других факторов. Трудно представить генетические модели, которые могли бы проверить эту гипотезу. Другие эксперименты не способны проверить, происходят ли специфические клеточные изменения именно через эти сигнальные механизмы, а не опосредованно. Степень, в которой такие свойства, как гомеостаз сигналинга АФК и организация структуры хроматина, являются ключевыми событиями в канцерогенезе, также остается неясной [61].
В будущем подобная специфичность и способность экспериментально проверить эти гипотезы может появиться из наблюдения количественных аспектов механизма, как это было в других исследованиях передачи внутриклеточных сигналов. Эксперименты, которые могут точно контролировать уровни ацетил-КоА и АФК, могут позволить исследователям выделить многие последствия эффекта Варбурга.
Заключение
Обширное изучение эффекта Варбурга и его функций в опухолевых клетках обеспечило понимание как его причин, так и условий для распространения раковых клеток [29,52]. И тем более удивительным является наличие пробелов в знаниях относительно истоков данного эффекта. Вероятно, потребуется лучшее понимание биологии эффекта Варбурга, чтобы достичь прогресса в терапии и профилактике рака, прибегая к нутритивному и фармакологическому воздействию на метаболизм, а также используя метаболизм глюкозы для управления иммунной системой, что в настоящее время является предметом пристального интереса учёных.
Вопросы, пока остающиеся без ответа:
- Как эффект Варбурга способствует развитию рака?
- Вследствие чего рост опухоли попадает в зависимость от эффекта Варбурга?
- Как разработать экспериментальную систему, которая позволила бы окончательно проверить все гипотезы относительно функций эффекта Варбурга?
- Имеет ли какая-либо из функций эффекта Варбурга непосредственное терапевтическое значение?
- Обеспечит ли знание функций эффекта Варбурга понимание его роли в эволюции опухолей?
- Являются ли необходимые условия для осуществления эффекта Варбурга ключом к пониманию его функций?
