
Нейрофизиология речи

Человеческий язык — пожалуй, самое гениальное, что когда-либо изобретала природа для передачи информации после генетического кода. Империя, созданная им за время существования нашей цивилизации, не уступает владениям генов: каждое явление человеческой культуры несет в себе язык и существует за счет него, как несет в себе геном и является его воплощением каждое живое существо.
Как и геном, язык обслуживает множество систем:
-
Мозг не только координирует воспроизведение и понимание речи — процессы мышления, насколько они нам доступны, даны в форме внутренней речи:
-
в восприятии речи участвуют первичные и вторичные корковые центры анализаторов, которые регистрируют соответствующий сигнал и квалифицируют как языковой;
-
в воспроизведении речи участвуют моторные и премоторные области, которые управляют движениями мышц, вовлеченных в речевую продукцию;
-
речь связана с эмоциональным регулированием: таким образом осуществляется контроль над интонацией, темпом и громкостью голоса, а также связь мимики и жестов с передаваемой словесно информацией — все это входит в понятие «невербальное общение»; невербальное сообщение воспринимается и интерпретируется одновременно с вербальным;
-
речь связана с памятью: смысловые единицы, зрительные и звуковые образы слов хранятся в памяти отдельно; все эти объемы информации интегрированы и внутренне организованы так, что доступ к ним занимает незначительное время;
-
речь связана с мышлением: современная позиция когнитивистики такова, что без речи мышление, в первую очередь абстрактно-логическое, невозможно [1].
-
-
Устная речь:
-
для воспроизведения необходимы координированные движения органов ротовой полости (относятся к пищеварительной системе) и мышц гортани; также важно размеренное дыхание (дыхательная система);
-
в восприятии участвует слуховой анализатор, хорошо приспособленный именно к тому диапазону громкости, в котором звучит человеческая речь.
-
-
Письменная речь:
-
для воспроизведения нужны сложные мелкие движения мышц руки, доведенные до автоматизма;
-
в восприятии участвует зрительный анализатор;
-
в распознавании текстов, написанных шрифтом Брайля, участвует тактильный анализатор.
-
Нам все это кажется естественным, но попробуем разобраться в грандиозной подспудной работе, которую при этом проделывает наш мозг.
С точки зрения врача, проблема речи — пограничная между неврологией и психиатрией. Речь задействует множество мозговых структур, и любое их нарушение, функциональное или органическое, с высокой вероятностью скажется на ней.
Около 40 % инсультов сопровождаются стойким или преходящим нарушением речи; именно его часто замечают первым — настолько этот синдром ясен и определен [2]. Но сохраняет ли такой пациент в полной мере мышление и собственную личность, которые «зашифрованы» в языке, мы не знаем, поскольку контакт с ним серьезно затруднен.
Для психиатра речь — «основной инструмент»: большинство симптомов и синдромов выявляются в ходе беседы, другого подхода к оценке мышления просто нет. В то же время о физиологических основах речи мы знаем больше, чем о других психических функциях, и в этом смысле она тоже могла бы послужить ключом к остальным психиатрическим симптомам.
Методы исследования речи
Ученых интересует локализация структур, контролирующих понимание и воспроизведение речи, связи между ними, этапы обработки речевого сигнала и роль каждой структуры на этих этапах.
Для подобных исследований необходимо создать контролируемую и как можно более простую речевую ситуацию (чтение или прослушивание слов или предложений, называние картинок), свести изменения в этой ситуации только к одному из аспектов речи (высота голоса, с которой произносится слово; соответствие слова контексту предложения) и получить данные о локализации происходящих в мозге изменений. Последнее достигается двумя способами: либо применяют функциональную нейровизуализацию и другие дополнительные методы исследования, позволяющие локализовать функционирующие в данный момент структуры, либо подбирают пациентов с поражением определенных областей и проводят детальное нейрокогнитивное исследование в сравнении со здоровыми людьми из группы контроля, определяя, какие функции при этом поражении выпадают. Если такие исследования дополнены функциональной нейровизуализацией, они представляют особый интерес еще и как отражение процессов нейрональной пластичности. Проблема всех исследований мозговой организации речи в том, что у одного пациента по данным разных методов исследования результаты могут существенно отличаться, и еще более существенные отличия получаются при сравнении разных пациентов; причиной этому может быть та же нейрональная пластичность [3].
Еще один принципиальный вопрос исследований речи — языки. Речь интересует нейрофизиологов как общее явление, но уйти от языковых различий невозможно даже на уровне экстраполяций на речь наших знаний о слуховом восприятии и движениях: еще в младенчестве внимание к звукам родной речи и звукам другого языка начинает различаться, хотя все остальные звуки одинаково воспринимаются детьми, воспитывающимися в разной языковой среде; ходить мы все учимся одинаково, но работа всех участвующих в речевой продукции мышц, вплоть до их тонуса и положения в покое, доводится до автоматизма в режиме, соответствующем конкретному языку [2]. Еще более сложный вопрос — исследование связи мышления, памяти с речью; установить эквивалентность «смыслов» на разных языках практически невозможно.
Морфологический метод исследования
Первые предположения о нейрофизиологии речи делались на основе наблюдений за пациентами, которые утратили способность говорить: подробно описав картину расстройства при жизни больного, после его смерти врач проводил вскрытие и соотносил патологоанатомические изменения с клиническими. Считаются классическими и иногда приводятся в учебниках в качестве исчерпывающего отчета о мозговой организации речи две работы:
-
за авторством Поля Брока — о поражении задней части нижней лобной извилины слева у пациента, повторявшего только слог «тан» в ответ на любой вопрос. Другие пациенты с такой локализацией поражения понимали обращенную к ним речь, но сами произносили только отдельные слова с большим трудом. Такой вид расстройства речи впоследствии стали называть моторной афазией, а соответствующую зону поражения — моторным центром речи [4]. Также Поль Брока отметил, что у большинства пациентов с нарушениями речи были патологические образования именно в левом полушарии — идея о функциональной асимметрии мозга принадлежит ему [5];
-
за авторством Карла Вернике — о поражении задней части верхней височной извилины у пациентов, которые не понимали обращенную к ним речь и сами изъяснялись непонятно, хотя и без затруднений, не осознавая собственных ошибок. Такое расстройство речи было названо сенсорной афазией [6].
Есть и отечественный ученый, чьи работы по изучению речи знают неврологи по всему миру — психолог Александр Романович Лурия. Во время Великой Отечественной войны он заведовал нейрохирургическим эвакогоспиталем, где и находил материал для работы, с той разницей, что данные о топике поражения знал из протоколов операций [7].
За время изучения проблемы речи накопилось немало таких клинических наблюдений: при повреждении каких структур развивается изолированный аграмматизм, диспросодия (нарушение ритмического построения речи с исчезновением из нее смысловых и синтаксических ударений, следствием чего является ее монотонность — прим. ред.); в учебнике неврологии они теряются в длинном списке терминов, называющих выпадение функций, и только особенно старательные студенты иногда выучивают парочку к экзамену. На начальных этапах такой фактический материал действительно интересен (как для студентов, так и для ученых), но со временем становится просто мертвым грузом. Сейчас, особенно когда неврологи принимают в расчет пластичность мозга и уходят от механической концепции «поражение области — выпадение ее функции», больший интерес представляют функциональные взаимоотношения, общие принципы, пути передачи возбуждения.
Электрофизиологические исследования
В первую очередь речь идет о методе вызванных потенциалов. Область, в которой генерируется выявленный потенциал, можно предположить, исходя из его выраженности в разных отведениях. В пример можно привести следующие важные исследования:
— потенциал N400, негативный, достигает максимальной амплитуды через 400 мс после предъявления соответствующего стимула, наиболее выражен в центропариетальной зоне, отражает процессы семантического анализа, то есть осознания смысла. Никакие изменения характеристик предлагаемого стимула (изменения размера букв, высоты голоса, которым произносились слова) не повлияли на амплитуду N400, он реагирует только на слова, резко выбивающиеся из контекста предложения (как в стихотворениях обэриутов, например, у Александра Введенского — «Больной, который стал волной») [8] или целого текста (например, когда рассказ о вегетарианце заканчивается словами «он заказал в ресторане стейк») [9];
— потенциал LAN (передняя левая негативная волна), по задержке напоминающий N400, но наиболее выраженный в отведениях от левых лобных отделов, реагирует на использование частей речи, не вписывающихся в синтаксическую структуру данного предложения (например, «мягко игрушка» вместо «мягкая игрушка») [10];
— потенциал p600 (синтаксический позитивный сдвиг) достигает максимальной амплитуды через 600 мс после предъявления стимула, наиболее выражен в отведениях от центропариетальной области. Стимулом при этом являются предложения синтаксически неправильные или со сложной структурой (например, двусмысленные газетные заголовки) [11].
Электростимуляционные исследования
Непосредственная электрическая стимуляция проводится во время операций на мозге с целью точного разграничения функциональных отделов и участков, подлежащих удалению. Пациенты находятся в сознании, и им могут быть предложены вышеперечисленные простые задания по распознаванию и произнесению слов. Хотя это непосредственный и точный метод изучения локализации речевых структур у отдельного пациента, его ограничение состоит в том, что практически единственный ответ на стимуляцию — speech arrest, невозможность речевой продукции. Таким образом, можно достаточно подробно описать, какие структуры в принципе вовлечены в речевую функцию, но как идет возбуждение и на каком этапе процесс был прерван раздражением в определенной точке исследователь показать не может.
Наиболее известное из подобных исследований принадлежит канадскому нейрохирургу Уайлдеру Пенфилду. Результаты своей работы он описал в монографии «Язык и мозговые механизмы», до сих пор актуальной для неврологов и специалистов по когнитивистике [12].
Сюда же можно отнести исследования с применением глубокой мозговой стимуляции [13] и транскраниальной магнитной стимуляции [14], которые начали проводить сравнительно недавно.
Методы функциональной нейровизуализации
На сегодняшний день являются ведущими методами при изучении нейрофизиологии речи. Позволяют судить о том, какие структуры функционируют в данный момент.
— функциональная магнитно-резонансная томография (фМРТ) и позитронно-эмиссионная томография исходят из прямых или косвенных данных об увеличении метаболизма в определенных областях мозга;
— диффузно-тензорная МРТ позволяет визуализировать проводящие пути головного мозга, связывающие эти области, путем расчета направлений диффузии воды;
— магнитоэнцефалография (МЭГ) дает возможность измерить и графически представить магнитные поля, которые образуются вследствие электрической активности мозга.
Структуры и проводящие пути
Классическая модель Вернике — Лихтгейма — Гешвинда
Согласно этой ранней модели, основанной на морфологических исследованиях [15], за воспроизведение речи отвечает структура на задней части нижней лобной извилины слева — зона Брока. На всем известном моторном гомункулусе эта область как раз соответствует мышцам лица, языка, гортани, участвующим в артикуляции и фонации. Здесь «хранятся» моторные образы слов — приобретенные в опыте речи приемы артикуляции, необходимые для произнесения и комбинации отдельных фонем.
Структура, ответственная за понимание речи — зона Вернике, задняя часть верхней височной извилины. Эта область примыкает к слуховой коре, специализирована для восприятия речи и содержит сенсорные образы слов — звуки и их комбинации, характерные для родного языка, которые ребенок может вычленять из всех окружающих звуков и запоминать с первых дней жизни.
Также модель утверждает, что ни моторные, ни сенсорные образы не несут смыслового наполнения — этот аспект речи не может быть конкретно локализован в мозге,но, тем не менее, в процессе обработки сигнала должно происходить обращение к «хранилищу» концептов — смысловых единиц нашей речи.
Поражение каждой из этих зон приводит к соответствующей речевой дисфункции, но для полноценной речи не менее важна их связь:
— Основной путь, связывающий зону Брока и зону Вернике — аркуатный пучок. При его поражении развивается кондукционная афазия: пациент способен понимать устную и письменную речь, но говорит с ошибками, которые осознает, но самостоятельно исправить не может. Спонтанная речь и повторение за другим человеком у таких пациентов затруднены;
— Также существуют связи между зонами Брока и Вернике и «хранилищем» концептов; разрушение этих связей приводит к так называемой транскортикальной афазии;
— Нарушение связи между зоной Вернике и областью, содержащей концепты слов, проявляется транскортикальной сенсорной афазией: пациент не понимает обращенную к нему речь и даже собственные слова, при этом свободно говорит с большим количеством ошибок (грамматические неточности он может исправить, если ему на них указать) и может без затруднений повторить любое высказывание;
— Нарушение связи между «хранилищем» концептов и зоной Брока проявляется транскортикальной моторной афазией: так же, как при моторной афазии, пациент говорит с большим трудом, односложно, аграмматично, но хорошо понимает обращенную к нему речь; отличие состоит в том, что при транскортикальной моторной афазии пациенты не способны повторить то, что слышат;
— Нарушение связи между первичной слуховой корой и зоной Вернике проявляется избирательной глухотой к словам: все остальные звуки такие пациенты слышат и правильно интерпретируют, но речь звучит для них как шум, расслышать в ней слова они не могут;
— Нарушение связей на любом этапе двигательного пути после зоны Брока — дизартрия, преимущественно моторное нарушение речи, никак не затрагивающее ее понимание.
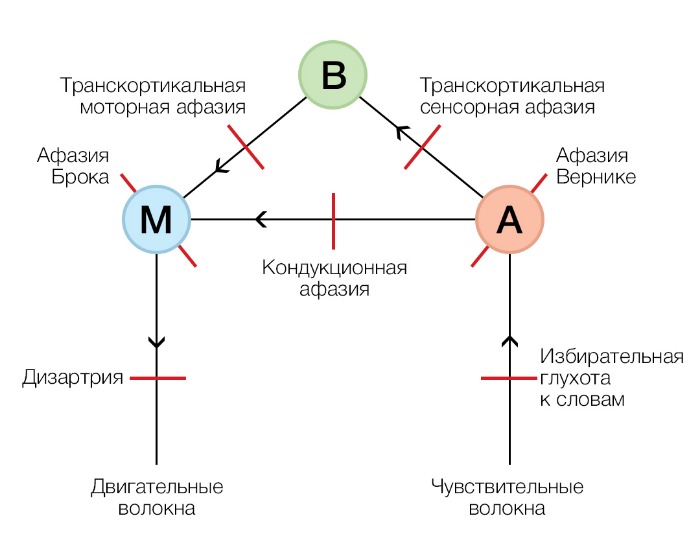
Проблема этой модели, во-первых, в том, что при дальнейших исследованиях пациентов с афазией, в том числе с использованием нейровизуализации уже на современном этапе, оказалось, что клиника афазии Брока (как и Вернике) не всегда, а в достаточно небольшом количестве случаев [16, 17] сопутствует поражению этой зоны, и наоборот. Во-вторых, классическая модель не разъясняет особенности обработки фонетического, лексического и синтаксического аспектов речи при ее восприятии и воспроизведении [18]. Такие термины, как «зона Брока», «зона Вернике» и соответствующие именные афазии неточны и для современной науки о языке имеют скорее историческое значение.
Современные представления
Модель «двойного потока»
Изначально похожая схема была открыта для зрительного восприятия: после обработки в зрительной коре затылочной доли сигнал следует вентральным и дорсальным потоками: вентральный идет к височной доле и необходим для опознавания объекта (отвечает на вопрос «что?»), а дорсальный идет к теменной доле и предназначен для распознавания пространственных характеристик объекта, его положения и возможных способов взаимодействия с ним (отвечает на вопрос «как?») [20].
Аналогично существуют два потока для звуков речи (рис. 2). Первичная обработка (касающаяся фонологического состава и частотно-временных характеристик звука) происходит в задней части верхней височной извилины и областях, окружающих верхнюю височную борозду (по большей части это и есть зона Вернике). Вентральный поток направляется к передним и средним отделам височной доли, он участвует в собственно распознавании и лексическом анализе речи (понимании ее смысла, соотнесении с хранящимися в памяти концептами). Дорсальный участвует в сенсомоторной интеграции, то есть соотнесении с моторными образами слов, и включает два отдельных пути: оба проходят через височно-теменную область, затем первый направляется к нижней лобной извилине, а второй — к префронтальной коре. Предположительно один из них участвует в восприятии и проекции на артикуляцию отдельных сегментов членораздельной речи, то есть отвечает за простейшие фонетические навыки, а второй — управляет построением последовательностей сегментов и обеспечивает более сложные навыки, такие как, например, обучение новым словам.
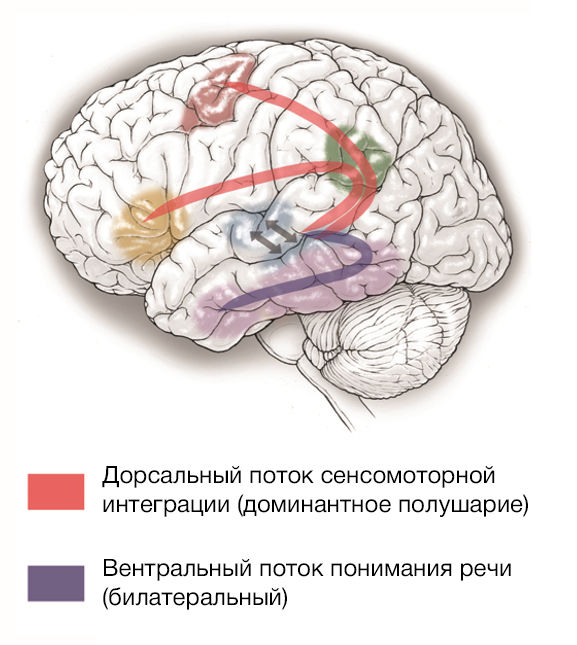
Есть варианты внутри этой модели:
-
модель Хикока-Попеля утверждает, что восприятие речи (через функционирование вентрального потока) происходит билатерально, а промежуточный центр дорсальных потоков (работающих исключительно в доминирующем полушарии) помещен в височно-теменную зону вдоль сильвиевой борозды, на стыке височной площадки (planum temporale) и теменной покрышки (operculum parietale), и этот центр, по их мнению, участвует только в сенсомоторной интеграции [21];
-
модель Раушекера-Скотта предполагает, что процессы по обработке воспринимаемой речи происходят только в доминантном речевом полушарии, а промежуточный центр дорсальных потоков располагается более медиально, в нижней части теменной доли, и кроме сенсомоторной интеграции он принимает участие в пространственной локализации звукового стимула [22].
Дополнения по данным диффузно-тензорной МРТ
Французский нейрохирург Гюг Дюффо первым провел объемную работу по интраоперационной трактографии у пациентов с афазией [27].
Основные пути, входящие в дорсальный поток — аркуатный пучок и верхний продольный пучок (superior longitudinal fasciculus, SLF). Верхний продольный пучок, в свою очередь, состоит из четырех компонентов:
- SLF I (не участвует в речевых процессах и далее не обсуждается);
- SLF II — соединяет угловую извилину (на стыке теменной, височной и затылочных долей, исторически входила в зону Вернике) с дорсальной премоторной и префронтальной корой [23, 26];
- SLF III — соединяет надкраевую извилину (в составе нижней теменной дольки) с вентральной префрональной корой [24, 26];
- SLF-tp — идет от нижней теменной дольки к задней теменной коре [25, 26, 27].
Интраоперационное раздражение SLF II и III приводит к расстройствам артикуляции (дизартриям) [24], а раздражение SLF III и SLF-tp — к ошибкам при повторении [28].
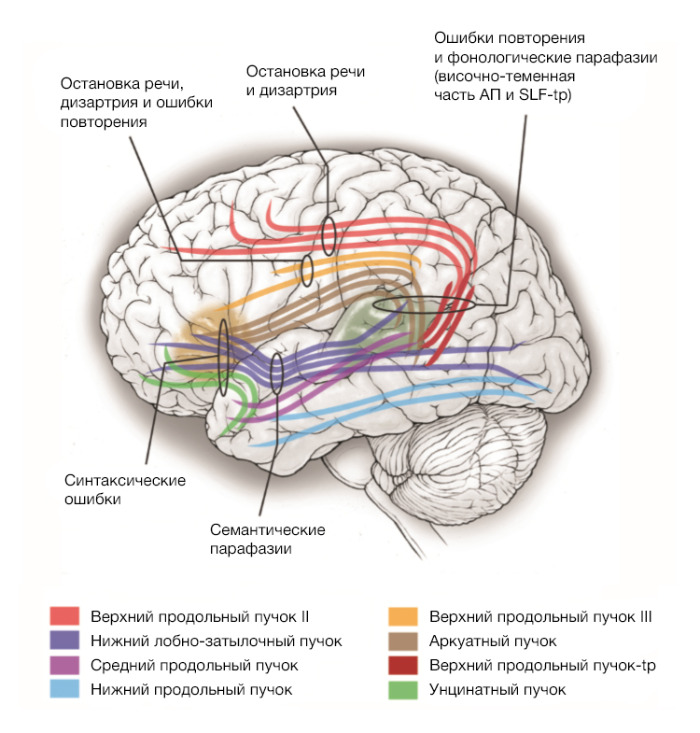
Аркуатный пучок — в классической модели единственный проводящий путь, — соединяет зону Брока с зоной Вернике. По данным трактографии, основная часть его волокон идет от покрышечной части лобной коры к задним областям височной, однако есть и ответвления. В лобной доле ответвления идут к покрышечной части нижней лобной извилины и вентральной премоторной коре [23, 29]. В височной части пучка основные ответвления идут к верхней и средней височной извилинам [30]. Сделанное на основе клинических и морфологических данных предположение, что нарушения проведения по аркуатному пучку ведут к кондукционной афазии, было подтверждено электростимуляционными исследованиями [31].
Сомнительным остается вопрос об отношении височно-теменного пути к аркуатному пучку. Имеются данные, что этот путь имеет значение в обработке звуковых сигналов и активируется при повторении за собеседником [32], хотя морфологически он достаточно отдален.
В вентральном потоке (участвующем в основном в семантической и синтаксической обработке) возбуждение также движется по нескольким путям.
Нижний лобно-затылочный пучок соединяет нижнюю лобную кору и дорсолатеральную фронтальную кору с задней частью височной и с теменной долей. Он проходит через переднюю ножку наружной капсулы в височную долю и дает ответвления к средней и нижней височным извилинам, а также затылочной доле [33]. При стимуляции этого пучка в эксперименте наблюдают семантические парафазии [34].
Данные по поводу унцинатного пучка, соединяющего переднюю часть височной доли с нижними отделами лобной, противоречивы. Хотя после его удаления пациенты не могут вспомнить имена знакомых [35], на данный момент признано, что его функция в распознавании смысла речи непостоянна [36].
Было показано, что способность людей выучить простое грамматическое правило задействует путь, соединяющий нижнюю лобную кору с задними отделами средней и верхней височных извилин, однако пока не определено, что это за путь — нижний лобно-затылочный пучок или унцинатный [37].
В речевом процессе может участвовать и подмозолистый пучок, проходящий между хвостатым ядром и поясной извилиной (или дополнительной моторной областью), а именно медиальная его часть. Если инсульт затрагивал эту область вместе со средней третью перивентрикулярного белого вещества, у пациентов развивалась тяжелая афазия с полной речевой аспонтанностью или речевыми стереотипиями, однако при поражении этих структур по отдельности существенных нарушений не наблюдали [38]. Электростимуляционные исследования подтверждают клинические данные: при интраоперационной стимуляции подмозолистого пучка получали транскортикальную моторную афазию, а при стимуляции перивентрикулярного белого вещества — дизартрию [27, 39]. Существует мнение, что в этой области концентрируются все проводящие пути от структур, участвующих в речевой продукции. Именно с этим связан, например, синдром дополнительной моторной области — преходящая потеря речи при опухолях этой зоны.
Недавно был описан также поперечный лобный путь, объединяющий верхнюю, среднюю и нижнюю лобные извилины [40]. Известно, что он дает выходы к стриарной системе, от него зависит вербальная беглость, а также он связан с такими нарушениями речи, как заикание [41].
Межполушарная асимметрия
Полушария мозга функционально и анатомически неравноценны, и каждый из нас хорошо знает это на собственном примере: большинство действий удобнее выполнять правой (меньшему количеству людей — левой) рукой, потому что отвечающее за эту сторону тела противоположное полушарие доминирует. Право- или леворукость — самый очевидный признак различия между полушариями, но на самом деле их гораздо больше: например, у доминирующего полушария больше связей с активирующими структурами ствола, а стимуляция симметричных точек инсулярной области с разных сторон может вызывать прямо противоположный вегетативный ответ [42]. И, хотя у здорового человека это никак внешне не проявляется, асимметрия существует также и в отношении функции речи. Более того, если расходятся результаты определения доминантного полушария по ведущей руке и по строению/активности речевых структур, решающими считаются последние. Причем левшей оказывается больше, чем людей, у которых латерализация речевой функции правосторонняя [43].
Леворуких гораздо меньше, поэтому их редко включают в исследования, а стараются скорее исключить для определенности результатов. К тому же, не только стороны, но и степени латерализации при направленном исследовании у них оказываются иными. Поэтому, говоря далее в этой статье о функции разных полушарий, будем держать в голове обычного правшу с доминантным левым полушарием.
Первое же исследование людей с «расщепленным мозгом», перенесших операцию по пересечению мозолистого тела, продемонстрировало, что речь при этом затрагивается, хотя не повреждается ни одна из структур, которые мы обсуждали ранее как участвующие в ее восприятии и продукции [44]. Такой пациент правильно называл предмет, который был представлен в правом зрительном поле, и, соответственно, воспринятая информация обрабатывалась зрительной корой левого полушария; предмет, представленный в левом зрительном поле (информация о котором обрабатывалась в коре правого полушария) пациент после каллозотомии назвать не мог и утверждал, что ничего не видел. Но когда условия эксперимента изменили и пациент должен был сигнализировать, что увидел вспышку света, с помощью азбуки Морзе, не используя речь, он справился с заданием: если предмет предъявляли в левом зрительном поле, он правильно подавал сигнал левой рукой, но все еще говорил, что ничего не увидел. Именно речевому описанию было доступно только то, что было обработано в левом полушарии, когда связи с правым пересекались, речевое выражение «его» информации становилось невозможным [2]. Но другая часть речевой функции — восприятие — у посткаллозотомических пациентов оказалось практически нормальной: все, что произносил экспериментатор, такие пациенты слышали и понимали достаточно хорошо, с какой бы стороны стимул ни предъявлялся [45]. На основании этих исследований предположили, что воспроизведение речи в большей степени латерализовано, чем понимание — что согласуется с моделью Хикока-Попеля.
Но у этой модели есть и сложности: хотя в экспериментах Зайделя [45] понимание отдельных слов сохранялось на удовлетворительном уровне, в более сложных тестах, направленных на выявление мягкой, незаметной на первый взгляд афазии у неврологических пациентов, перенесшие каллозотомию показывали плохие результаты, если стимул предъявлялся так, что его обрабатывало недоминантное полушарие. С этими данными согласуется предположение о несколько ином функционировании дорсального и вентрального путей, которое выдвинули авторы, изучавшие патологии речевого развития у детей. Суть его в том, что вентральный поток, хотя и функционирует билатерально, отвечает за распознавание только базовых синтаксических конструкций: коротких предложений, простых времен. Именно он на каком-то этапе эволюционного развития человеческой речи мог обеспечивать так называемый «протоязык». Дорсальный поток, в свою очередь, имеет отношение к расширенному синтаксису и является эволюционно более поздним и совершенным [46].
Все эти построения опровергают данные нейровизуализации: они однозначно показывают, что при восприятии речи активация заинтересованных структур асимметрична в той же степени, что и при воспроизведении [47, 48, 49]. Различия обнаружены в активации теменных и лобных центров: показано, что для последних характерна большая степень латерализации, это косвенно свидетельствует в пользу возможной билатеральной активности вентрального потока, которую, однако, по мере повышения уровня обработки до более сложного «перебивает» более латерализованный дорсальный [50]. Однако последнее объемное исследование сходного дизайна не подтверждает такого распределения, хотя поднимает ряд других интересных вопросов: например, как полушария действуют друг на друга? Между симметричными речевыми структурами разных полушарий возможно существует что-то вроде конкуренции, или, наоборот, взаимной активации. То, что мы наблюдаем у посткаллозотомических пациентов, может быть результатом утраты этих влияний и разбалансировки, а у здоровых пациентов — их равновесия [51].
Речь и эмоции
Латерализация речевых функций — для ученых вопрос до сих пор темный, но если спросить обывателя, в чем разница между полушариями, он ответит, что левое полушарие скучное и логичное, а правое — эмоциональное и творческое. В том самом эксперименте с расщепленным мозгом было еще много этапов, кроме называния предметов и понимания слов; выяснилось, что, хотя с речью правое полушарие полноценно не справляется, есть задачи, которые остались на его долю: например, конкретно-образное мышление, задания, связанные с манипуляцией предметами, изображениями, а не абстрактными понятиями [2]. Что касается речи, пациент с повреждением правого полушария к словесному общению способен, адекватен и понятен собеседнику, но теряет все невербальные аспекты: его речь монотонна и лишена всякой эмоциональной составляющей, воспринимать их он также не может. Образные выражения, конкретные описания — все, что направлено на соответствующий отклик у собеседника, также становится ему недоступно [52]. Такова точка зрения классической неврологии.
По всей видимости, межполушарная асимметрия в нейрофизиологическом обеспечении эмоций существует и вне связи с речью, только здесь для разной модальности эмоций характерна право- или левосторонняя латерализация [53], что было известно еще из клинических исследований [52]. Эмоциональные компоненты именно речи, какой бы модальности ни были эмоции, традиционно приписывают активности одного, правого полушария, однако здесь все не так однозначно.
Абсолютно все модуляции голоса в процессе воспроизведения речи — темп, громкость, тон, ритм, тембр — объединяются термином просодия. Различают просодию лингвистическую, характерную для определенного языка в целом, которой приходится учиться так же, как чужой грамматике, чтобы речь звучала естественно; и эмоциональную, которая подчиняется гораздо более общим принципам и каждому здоровому человеку врождена. Теоретически, лингвистическая просодия теснее связана с синтаксическими правилами языка и, наравне с их использованием, будет искажаться при поражении доминантного полушария.
По результатам последнего мета-анализа, исследующего восприятие просодии, данные в пользу правосторонней латерализации эмоциональной просодии есть, но они оказались статистически малозначимыми; предположение о левосторонней латерализации лингвистической просодии и вовсе не подтвердилось. В то же время, исследования показывают, что обе разновидности просодии искажаются при поражении любого полушария [54]. То есть, как и в случае с восприятием и воспроизведением речи, который обсуждался выше, говорить о четкой функциональной специализации полушарий нельзя, имеет место именно их взаимодействие.
Объяснить преимущественно билатеральное распределение просодии может гипотеза, опирающаяся на акустические характеристики речи: логично, что мозг ориентируется на физические показатели звука, потому что искусственных определений «эмоциональная», «лингвистическая» просодия для нейронов не существует. Согласно этой гипотезе, левое полушарие более специализировано для распознавания временных характеристик, а правое — частотных [55]. Именно такое распределение было показано для передней части верхней височной извилины, где информация обрабатывается сразу после первичной слуховой коры [57]. Но если бы это действительно вело в конечном итоге к определению эмоций, на последнем этапе обработки активировались бы те же зоны, что и при восприятии этой эмоции другими путями (при рассматривании картинок, например), а такой корреляции уже не обнаружили [57]. Тем не менее, эту теорию пока нет оснований сбрасывать со счетов — как и концепцию правосторонней латерализации просодических характеристик. Последняя подтверждается, например, недавним трактографическим анализом [58].
Основываясь на мнении, что правосторонняя латерализация все же имеет место, полагают, что и восприятие, и воспроизведение просодической речи обеспечены теми же нейронными путями, что описаны для вербальной коммуникации слева, просто в правом полушарии вентральный и дорсальный поток специализированы для других целей [59].
Мы уже говорили о том, что дорсальный поток связывает услышанное с моторными образами, и таким способом участвует в восприятии речи. Поэтому детям удобнее читать вслух — одновременная артикуляция улучшает понимание [60, 61]. При изучении речи в первую очередь привлекает внимание исследователя активация в области нижней лобной и верхней височной извилин. Но, изучая восприятие, можно также отметить активацию первичной моторной коры, которая никакими мышечными сокращениями не сопровождается. Поскольку первичная двигательная кора изучена достаточно хорошо, мы можем определить, какими мышцами обычно «управляет» активный сейчас участок — именно так и обнаружили, что, слыша слова, моторная кора «вспоминает», как бы она распорядилась органами артикуляции, если бы пришлось их произносить. Аналогичная активация моторной коры была обнаружена при изучении просодии — в зонах, которые отвечают за напряжение голосовых связок, то есть вокализацию. Чтобы представить себе эмоцию другого человека, моторная кора «представляет себе», как должны сработать наши собственные связки, воспроизводя эту интонацию [62, 63].
Из структур вентрального пути справа активны в заданиях с распознаванием эмоциональной речи две: области, окружающие задние и передние отделы верхней височной борозды. Предположительно, задние отделы ответственны за обработку услышанной фразы именно как комплекса звуков, со всеми особенностями; в этом процессе нет несущественных деталей, в общую картину входит абсолютно все: особенности конкретного собеседника, данной фразы — все воспринимается, как есть. Роль передних отделов — выделить в услышанном звуковом фрагменте какую-то ведущую «мелодию», не зависящую от собеседника, которая уже известна, как несущая определенный смысл [64]. Именно эту струну затрагивает в нас музыка: хотя она может и не содержать слов, которые для нас более привычны в качестве средства коммуникации, музыка все же что-то нам «говорит», мы всегда можем назвать ее эмоцию, потому что путь ее восприятия аналогичен таковому для речевой просодии (включая иногда острое желание запеть, которое обеспечивает дорсальный поток) [65]. Другими словами, вентральный путь правого полушария — путь упрощения, абстрагирования звукового сообщения от всех данных, присущих конкретной ситуации (у задних отделов верхней височной борозды) к так называемому «просодическому Гештальту», целостному интонационному образу (у передних ее отделов) [56].
Кроме структур височной области, эмоциональную составляющую речи обеспечивают образования лимбической системы; наиболее достоверно показана роль базальных ядер [54]. Какое отношение они имеют к восприятию просодии, пока не выяснено, зато они непосредственно участвуют в двигательной организации речи.
Речь и память: ментальный лексикон
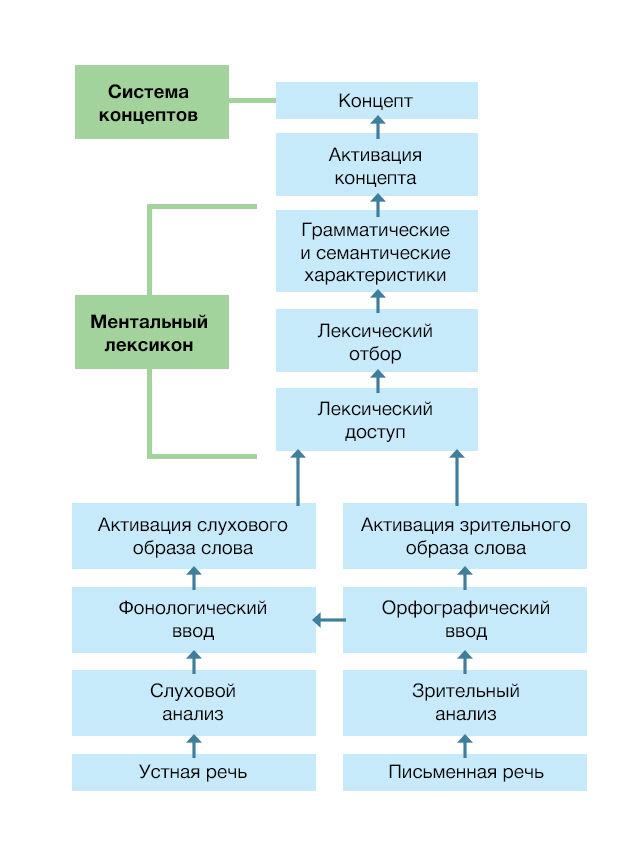
Прочитали мы слово, услышали или увидели предмет, который оно обозначает, или хотим донести до собеседника свою мысль — все пути ведут к хранящемуся в нашей памяти смыслу. Как именно эти смыслы хранятся, в какой доле, у какой борозды, мы сказать не можем. Мы не до конца понимаем, как работает речь даже и на более «приземленных» этапах, хорошо знакомых неврологам; по большому счету она остается для нас «черным ящиком», и что происходит внутри — загадка. Когда мы видим, что цепь прервалась, и пациент демонстрирует непонимание смысла, мы все еще не можем с уверенностью судить, на каком этапе это произошло — просто потому что не знаем всех этапов. Исчезли из памяти сами смыслы? Нарушился доступ к их хранилищу? Изменились связи между концептами, и теперь функционируют не так, как мы привыкли видеть? Поэтому про самую сердцевину речи, ее содержание, мы не можем сказать практически ничего определенного — но есть предположения, как понимание смысла речи организовано в головном мозге.
Ведь если задуматься, вычислительные возможности современных компьютеров гораздо больше, чем у мозга: создававшая его эволюция, в отличие от инженеров, никогда не начинала с нуля, располагая практически любыми материалами, и далеко не всегда исходила из принципа наибольшей эффективности. Но несмотря на множество ограничений, все необходимое для обмена информацией мозг выполняет с удивительной скоростью. Дело не в высоких скоростях проведения импульса или обменных процессах — это возможно благодаря эффективной организации всего, что мы помним: мозг — плохой компьютер, но хороший библиотекарь. Поэтому наиболее эффективны не те способы запоминания или улучшения памяти, которые повышают «технические» возможности мозга (например, зубрежка), а те, которые направлены на систематизацию информации.
Ментальный лексикон — это совокупность информации о словах, которая включает собственно значение слова (семантический компонент), все роли, которое оно может играть в предложении (синтаксический компонент), а также воспринимаемые и воспроизводимые формы слов (как они звучат или выглядят на письме, какие мышцы необходимо привести в движение, чтобы этого звучания/изображения добиться) [2]. Он является одновременно конечной точкой восприятия речи и начальной — воспроизведения; как в кладовой, в ментальном лексиконе мозг отыскивает и узнает то, что говорят нам, а с другой стороны — выбирает то, что мы сами собираемся сказать.
Как и в случае любых движений и любых видов восприятия, затруднения в поиске нейрональных коррелят появляются, как только мы пытаемся отойти от периферических отделов системы и ее первичных корковых областей, заглянуть глубже, туда, где по сути и происходят наши мыслительные процессы. Здесь и наука о речи переходит в область гипотез: процессы, ведущие к тому, что мы получаем на выходе, возможно, устроены именно так, но физиологических коррелят, пошаговых путей от одной группы нейронов к другой у нас нет.
Пассивный словарный запас среднестатистического взрослого человека — в среднем около 50 тысяч слов [66]; в когнитивном тесте MOCA нормой считается назвать за 60 секунд 11 слов, начинающихся на одну и ту же букву [67] — из такого большого объема извлечь нужное слова мы можем достаточно быстро. Словари обычно располагают слова последовательно и в алфавитном порядке, и в поиске какого-то из них мы находим нужную букву, а потом просматриваем их одно за другим — способ, мягко говоря, небыстрый. Ментальный лексикон устроен по-другому [2, 68]:
Во-первых, его наименьшей единицей считаются скорее не отдельные слова, а морфемы — минимальные смыслосодержащие единицы [2]. Это корни, суффиксы, приставки. В то же время, у ментального лексикона может быть несколько уровней, и на одном из них минимальной единицей будет являться слово.
Во-вторых, он представляет собой не список, который последовательно просматривается, а сеть, где каждая единица связана со множеством других [69]. Суть такой организации в том, что слова, связанные по смыслу, часто употребляются в одной речевой ситуации, и если речь заходит об одном, разумно было бы «держать под рукой» и остальные. Чем чаще слова используются вместе, тем сильнее будет связь между ними, тем больше наклонность переходить именно по этой связи в ходе ассоциативного процесса.
Исходя из этого, изобретенный Фрейдом метод свободных ассоциаций обоснован: он действительно может обрисовать нам «психическую ситуацию» пациента, потому что теснота связей абсолютно индивидуальна. Работая с неврозами, Фрейд рассчитывал эту связь понять, но у психотических пациентов ассоциативный процесс зачастую нарушен так, что проследить ее мы просто не можем; выходит, именно в особенностях работы ментального лексикона можно найти смысл психоза. Что заставляет ассоциативный процесс пойти по настолько нестандартному пути, вдруг свернуть к чему-то совершенно необъяснимому: перестройка связей или постоянная активность именно «бредовой» части лексикона, которую мозг решил почему-то все время «держать под рукой»?
В-третьих, у ментального лексикона не один организующий принцип (как у словаря — алфавит), а сразу несколько.
Ведущий — семантический, то есть связи осуществляются по значению (рис. 4) Важность этого принципа доказывает семантический прайминг-эффект (в русскоязычной литературе также используется термин «эффект предшествования»). В эксперименте используют два слова: «предшествующее» — это всегда реальное слово, и «целевое» — это может быть также настоящее слово, псевдослово, которое построено с соблюдением фонетических принципов, но отсутствует в языке, или просто нечитаемый набор букв. Испытуемому нужно, нажав на кнопку, решить, является ли показанное ему целевое слово настоящим, псевдословом (похоже по структуре на настоящее, но несуществующее, как «глокая куздра») или непроизносимым набором букв — так называемая ситуация «лексического решения». Если «предшествующее» слово связано по смыслу с «целевым», испытуемый выполняет задание в разы быстрее: память уже перешла к «разделу каталога», в котором находилось это слово в одной смысловой связке, оставалось только найти его [2, 70]
С другой стороны, учитывается также звуковой состав слова [2, 71]. Минимальная единица звукового состава — фонема; она не несет смысла сама по себе, но если заменить в слове одну фонему, изменится и его смысл: например, слова «кошка» и «мошка» отличаются одной фонемой, а по смыслу совершенно различны. Слова с похожим звучанием так же образуют сеть, как и слова, связанные по смыслу (предметы одного типа, одного цвета, объединенные одним действием), поэтому мы тяжелее отличаем их друг от друга, чем слова, состоящие из абсолютно разных фонем [72].
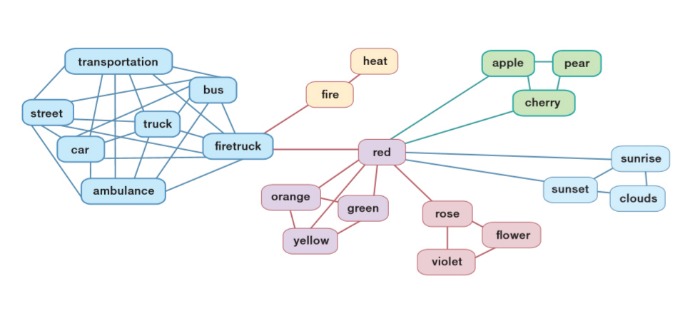
Ментальный лексикон — центральное звено организации речи. Однако есть мнение, что для восприятия и продукции слов существуют два отдельных лексикона [2].
Еще одна важная идея — концепция лексического доступа (рис. 5) [73]. Разговор о восприятии речи, насколько оно доступно неврологам, заканчивается в тот момент, когда звук квалифицируется как речевой и делится на фрагменты, по которым можно «отправить запрос» в библиотеку ментального лексикона. Если более ранние этапы мы можем локализовать в мозге, то относительно последующих предполагается только, что обработка происходит в три этапа:
-
Собственно лексический доступ — соотнесение полученного звукового (или зрительного — в памяти хранятся все разновидности, которым индивид был обучен) образа с образами слов, хранящимися в памяти. Каждое слово присутствует в «библиотеке» во всех грамматических формах (например, времена глаголов или роды прилагательных), за счет этого на этапе лексического доступа из звукового образа извлекается одновременно семантическая (по значению слова) и синтаксическая (по его форме) информация. Все похожие на услышанное (или увиденное) образы «активируются» и готовы к использованию в ближайшее время, например, при назывании слова «красный» активируются связанные с ним в семантическую сеть названия других цветов, предметы красного цвета и т. п.;
-
Лексический отбор — этап, на котором среди всех активированных словоформ отбирается наиболее подходящая к услышанному или увиденному;
-
Лексическая интеграция — объединение всех слов в более крупные структуры: словосочетания, предложения, вплоть до целых внутренне связных текстов.
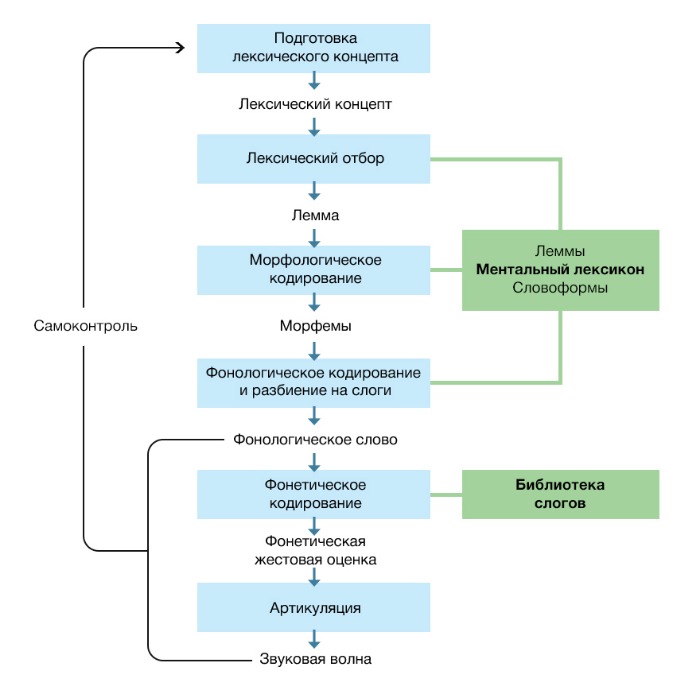
Тот же ученый предложил и концепцию формирования высказывания при воспроизводстве речи, по большому счету, схожую с концепцией лексического доступа при восприятии (рис. 6) [74].
Подготовка сообщения, которая включает макро- и микропланирование.
Коммуникация, по большому счету, — один из способов повлиять на окружающую среду. Поэтому коммуникация, как и любое действие, подчинена мотивационной сфере и может быть направлена на удовлетворение потребности или устранение неудобств. На примере инструктивных выражений это видно особенно отчетливо, но эту же цель, по всей видимости, имеют и другие наши высказывания, потому что иных источников мотивации просто нет. Таким образом, любое высказывание должно быть сформулировано исходя из результата, который мы хотим получить — этот аспект подготовки сообщения называется макропланированием.
При микропланировании разрабатывается вопрос, как информация должна быть представлена: какой группы слова использовать, в какие отношения их поставить.
После подготовки сообщения на выходе получается «концепт», общий смысл целого предложения.
Например, в эксперименте человеку показывают картинку со стадом коз, и он должен ее назвать ( это «макроплан»). «Микроплан» вызывает в памяти тот фрагмент сети смыслов, который может быть связан с этим изображением: кроме слова «коза», могут «активироваться», например, «овца», «рога», «деревня», «сыр» и так далее. Обратим внимание, что речь идет о смыслах, «концептах», а не о фрагменте ментального лексикона. Как говорилось выше, где в мозге хранится «смысл», то, что мы называем целостным пониманием, гештальтом, нам неизвестно — как и есть ли он вообще. Возможно, ощущение, которое мы называем «пониманием», «осознанием», работает как-то иначе. Ментальный лексикон — понятие гораздо более конкретное, и он распоряжается меньшими, более формальными единицами, чем «хранилище концептов». Точно так же носителями идей, действительно понимающими их, могут быть только люди, и, хотя все богатое разнообразие выражений для этих идей хранится в библиотеке, «одушевляют» книги только читающие их.
Формулирование — придание предложению конкретной формы, грамматической и фонетической, готовой к произнесению.
Концепт, полученный на первом этапе, взаимодействует уже с ментальным лексиконом; первое, с чем необходимо определиться — часть речи (в нашем случае поиск обратится к существительным). На выходе получаем соответствующую лемму — начальную форму слова. В нашем случае это именительный падеж существительного «коза».
С леммой связано множество разных словоформ — в нашем случае, существительное «коза» в разных числах и падежах. Нам нужно существительное именно во множественном числе — по этому принципу происходит лексический отбор словоформ.
Далее происходит морфемный анализ: лемма «коза» должна видоизмениться, чтобы соответствовать необходимому множественному числу: меняется словообразующая морфема, окончание с «а» на «ы».
Полученное слово нужно будет назвать, подготовкой к чему становится фонологический анализ: разбор количества слогов и положения ударения в слове. Информация, которую мы получаем на выходе — разложение слова по слогам, один из которых потребует изменения тона голоса, поскольку является ударным, — достаточно проста, чтобы «передать» дальше для формирования двигательной программы. Иногда в момент фонологического анализа процесс прерывается, тогда и возникает «феномен кончика языка»: мы прекрасно понимаем, о каком существительном идет речь, оно само не забыто, — но не можем его назвать.
Нейробиологических данных, которые затрагивали бы вопрос ментального лексикона, немного; они не позволяют составить целостную картину, но все же они есть.
Элизабет Варрингтон с коллегами исследовала проблему избирательных, (относящихся только к одной категории предметов) агнозий, и как они могут соотноситься с семантической памятью [75]. Выяснилось, что, как и гнозис, семантическая память разделена на «категории», для которых возможно также и ее изолированное нарушение. В неврологии привычное явление, что пациент не лишается функции полностью, если ее могут выполнять какие-то дополнительные пути или соседние, связанные структуры. Раз возможна такая избирательная амнезия, «выключающая» определенную «область смыслов», можно предположить, что в нашем ментальном лексиконе (или хранилище концептов?) далеко не каждое слово связано с каждым так, чтобы можно было прийти к нему другим путем; какое-то деление, рассредоточение все же есть. Варрингтон выделила такие группы: живые существа, которые в основном связаны с терминами, описывающими их внешний вид и другие свойства, воспринимаемые органами чувств; и искусственные предметы — они обычно описываются по функции, которую выполняют.
Больше того, для пациентов, которые затруднялись вспомнить слова, связанные с живыми существами, была характерна определенная локализация поражения — нижние и медиальные области височной коры, чаще в передних отделах. Передние отделы нижневисочной коры связаны со зрительным восприятием объекта (что как раз и дает нам информацию об их внешнем виде); тот самый «вентральный поток» для зрительного восприятия, ассоциацию с которым потом перенесли на речь, заканчивается именно здесь, и по итогам обработки на этом пути мы можем ответить на вопрос «что это такое?». Медиальные отделы связаны с гиппокампом и участвуют в извлечении информации из долговременной памяти.
О локализации поражений у пациентов с другим типом дефицита семантической памяти, касающегося предметов, созданных человеком, известно меньше. Однако немногие существующие исследования на этот счет говорят о вовлечении теменных и лобных областей слева, как раз тех, которые отвечают за пространственную организацию и планирование движений (отвечают на вопрос «как?»).
Концепцию Варрингтон не все поддержали: некоторые ученые утверждали, что ее нельзя принять из-за качества контрольных материалов, которые она использовала. Карамацца и Шелтон предложили собственную гипотезу: два отдела семантической памяти отвечают за одушевленные и неодушевленные предметы [76]. На практике категории, предложенные Варрингтон и ее оппонентами, пересекаются, но авторы настаивают на эволюционной обоснованности и психологической важности именно второго варианта: происхождение объекта — был он рожден или создан человеком — не так важно как то, приписываем ли мы ему одушевленность, свободную волю или нет. Однако ведущей на данный момент остается концепция Элизабет Варрингтон, и в дальнейшем исследователи пользовались ей. Выполненные позже и уже на здоровых пациентах нейровизуализационные исследования в целом ее подтвердили [77, 78].
Воспроизведение и восприятие речи
Продукция
Двигательный компонент организации речи всегда считался достаточно доступным для неврологов: по большом счету, движения, которые необходимы для воспроизведения звуков речи, принципиально не отличаются от движений другой скелетной мускулатуры.
Периферический речевой аппарат состоит из трех компонентов: дыхательного, голосового и артикуляционного [79].
Дыхательный компонент включает легкие и дыхательные пути. Если сравнить речь с течением реки, то голосовой и артикуляционный компонент — это берега, а дыхание — это вода, сама основа речи. Контроль дыхания преимущественно автономный: дыхательный центр находится в стволе, обладает собственным ритмом активности и управляется, если говорить упрощенно, по механизму обратной связи — в зависимости от кислотно-основного состояния крови и парциального давления газов в ней. «Точка приложения», за счет которой осуществляется управление, — дыхательные мышцы, однако большая их часть может сокращаться также и произвольно [80]. На протяжении эволюции человека способность к произвольному контролю дыхательной мускулатуры возрастала — это считают одним из факторов становления членораздельной речи [81]. Дыхание во время речи особым образом изменяется: поскольку сами слова произносятся на длинном выдохе, он занимает большую часть дыхательного цикла, вдох же наоборот становится глубоким и коротким. Объем вдыхаемого воздуха увеличивается примерно в три раза. Ритм дыхания также перестраивается: вдохи удачно приходятся на паузы между словами и предложениями [82, 83]. По всей видимости, моторные центры речи могут действовать также и на нейроны, регулирующие паттерн дыхания [84], но как конкретно это происходит у человека, до сих пор неизвестно. В экспериментах на животных было обнаружено стволовое ядро, которое координировало вокализацию и дыхание — вентролатеральное парабрахиальное; предполагают, что похожая структура есть и у человека [85].
Тонкости работы голосового и артикуляционного аппарата достойны отдельной статьи; скажем только, что для извлечения правильного звука необходимы определенные положения каждой из их частей. Все органы, необходимые для вокализации и артикуляции, — органы головы и шеи; они иннервируются черепно-мозговыми нервами, к двигательным ядрам которых подходят корково-ядерные пути от соответствующих отделов коры. Соматотопическая проекция мышц языка, гортани, жевательных и мимических мышц в двигательной коре занимает наиболее латеральные отделы прецентральной извилины — это и есть «зона Брока». Так же, как и для всех движений, подлежащих научению, — ходьбы, письма, игры в теннис, — для произношения отдельных букв, слов и предложений существуют двигательные программы, которые в определенной степени выполняются автоматически. Отбор и целенаправленное выполнение этих двигательных программ — задача экстрапирамидной системы [52]. Обязательный этап речевой продукции, как и любого движения — обратная связь: проприоцептивная, зрительная или слуховая [2].
Звучащая речь была первой системой, подходящей для обмена действительно сложными, детальными информационными сообщениями (хотя есть ученые, которые утверждают, что первична в ходе эволюции была жестовая речь [86]). Однако мы активно пользуемся и другими: например, той же жестовой речью, но наиболее часто — письмом. Здесь эволюция тоже не стоит на месте: письмо от руки, требующее разнообразных тонких движений кисти, сменяется печатанием: сначала на большой компьютерной клавиатуре, двумя руками, в стационарной позе за столом, а потом и на экране телефона, в маршрутке, одним большим пальцем. Все эти движения, технически абсолютно разные, привязываются у нас в голове к одной знаковой системе, но собственно исполнение требует вовлечения несколько иных структур, чем при разговоре.
Коротко остановимся на письме от руки: проверка почерка, в отличие от способности пользоваться мессенджерами, на сегодняшний день входит в арсенал невролога (возможно, это дело времени). Концепция формирования высказывания для написания начинает отличаться устной речи после морфемного анализа; в случае устного высказывания это был бы фонетический анализ и непосредственная проекция его результатов на двигательные программы. Здесь же нужен анализ графемический, то есть проекция на знаки письменности. Как уже говорилось, возможно, ментальный лексикон содержит сразу несколько параллельных сетей, и содержащую письменные образы слов в том числе. Однако письмо — умение, которое приходится специально осваивать, постоянно отсылая учащегося к опыту устной речи, поэтому более распространено мнение, что при построении сообщения для письма происходит фонетически-графемический переход, то есть в знаки письменности фраза «переводится» не сразу, а из соответствующих фонем [87]. Что касается собственно движений, в исследованиях нужно учитывать их уровневую организацию: с одной стороны, общий контур движения, который необходим, чтобы изобразить букву (он не привязан к определенной конечности или ее сегменту), с другой — статическая работа по сохранению необходимого положения руки и удержанию ручки [88]. Этим письменная речь отличается от устной: моторные программы для артикуляции «жесткие», а для письменной — достаточно легко проецируются на другие конечности.
По результатам метаанализа, который учитывал эту особенность, необходимы для письменной речи следующие три области:
-
верхнелобная — включает среднюю лобную извилину, область вокруг верхней лобной борозды;
-
верхнетеменная — нижняя и верхняя теменная долька, внутритеменная борозда;
-
первичная моторная и сенсомоторная кора.
Как уже говорилось, письмо от руки — достаточно строго латерализованная функция, и активация этих областей наблюдается в основном в правом полушарии.
Движения невозможны без многих других структур, например, экстрапирамидной системы, мозжечка, но, по всей видимости, именно эти области имеют отношение к созданию и воплощению «общего контура движения», который является сутью письменной речи.
Восприятие
Этот процесс устроен несколько сложнее. Разная высота и тембр голоса, шепот и крик, полный рот горячей картошки, вологодское оканье, — если вслушаться, нет двух людей, которые произносили бы одно и то же слово одинаково. Изучая перцепцию речи, ученые пытаются разбить ее на мельчайшие звуки и определить их физические характеристики, на которые мозг мог бы опираться, но они оказываются слишком разными и к тому же произносятся слитно, с довольно большой скоростью. То же касается и всех остальных видов речи: если начинать с мельчайших характеристик, двух похожих знаков не найдется. Но даже если нам показать текст, например, на японском языке, которым мы не владеем, мы сможем понять, что какие-то из этих символов одинаковые, какие-то чаще встречаются в начале предложения и так далее — как Шерлок Холмс в «Пляшущих человечках». Видеть эту однотипность, представлена ли она в звуках или в письменной информации, значит владеть самим принципом языка.
Устная речь
Любой звук, который мы слышим, сначала обрабатывается в первичной слуховой коре — в извилине Гешля. Она активируется всегда билатерально в ответ на любой звуковой раздражитель, но ассоциативные области слуховой коры, составляющие вентральный путь, активируются, только если мы слышим речь. Исследуя области, более и менее чувствительные к звукам речи в первичной и ассоциативной коре, Джеффри Биндер также показал, что семантические аспекты в этих областях не учитываются [89]. Недавние нейровизуализационные исследования показали, что направление обработки информации, которое устанавливает речевую природу звука — это и есть вентральный поток [90]. При этом различение слов и псевдослов на слух происходит очень быстро — в течение 50–80 миллисекунд [91].
Письменная речь
Навык чтения, как и восприятие устной речи, начинается с малого: следование горизонтальной или вертикальной линии, распознавание простых форм.
Так же, как и в случае слуховой коры, у зрительной есть ассоциативные области, которые более чувствительны к языковым символам, хотя сначала информация обрабатывается в первичной зрительной коре, чувствительной к любой зрительной стимуляции; они располагаются на стыке с височной корой [92]. Та же область коры отвечает за узнавание любых зрительных образов, обладающих определенной долей общности, например, лиц. Ученые попытались выделить регионы, которые были бы активны при просмотре именно символов языка; действительно, нейровизуализация показала,что есть очаги, активность которых очень высока при предъявлении фотографий, и на буквы они не реагировали; есть очаги, с которыми все было наоборот — высокая активность наблюдалась при предъявлении набора букв. Только в ответ на предъявление слова в этих самых областях никакого эффекта не увидели [93, 94]. Впоследствии исследование с использованием одновременно фМРТ и вызванных потенциалов выявило область, которая избирательно активировалась в ответ на настоящие слова, она расположена кпереди и латерально от V4 (область зрительной коры), это участок затылочно-височной борозды, расположенный слева [95] — то есть в восприятии письменной речи предположительно существует латерализация. Поражения именно этого участка ассоциированы с алексией — невозможностью читать, — однако со значением слов он никак не связан [96].
Заключение
Язык доступен нам в первую очередь как часть гуманитарной науки, именно так мы учим его в школе: члены предложения, части речи, фонетические разборы.
Но «морфем», «фонем» и «слов» в мозге нет — все это лингвистические термины, а мозг состоит из нейронов, образующих взаимно связанные функциональные группы. Задача биологической науки о речи — перевести лингвистические термины на язык нейронов, и в процессе решения этой задачи изменяется облик и лингвистики, и нейрофизиологии. В поисках нейрональных коррелят ученые опираются на лингвистическую теорию, но обнаруженное может ее изменить; с другой стороны, именно свойства мозга определяют возможность речи, такой, какой мы ее знаем, и именно нейробиология находит то общее, к чему лингвистика не всегда может свести языки мира.
Взрослым людям задают какие-то глупые задачки, просят отличить знакомое любому пятилетнему ребенку слово от беспорядочного набора букв — и на самом деле, ученые только играют с камешками на берегу океана. Самая интересная загадка — не разница между «кошкой» и «мошкой», а речь в принципе, одновременно явление нашей культуры и жизненно необходимая функция, такая же, как жевать или бегать; она может уходить корнями глубже, чем мы себе представляем.
По сравнению с другими философскими вопросами, биологи — и врачи вместе с ними, — взялись за язык совсем недавно, поэтому главных открытий от них мы еще только ждем. В конце концов, как говорил один польский писатель, язык — слишком важная вещь, чтобы доверить ее языковедам.
1. Чуприкова Н. И. Дифференционно-интеграционная теория развития как основа решения проблемы соотношения языка и мышления //Дифференционно-интеграционная теория развития.—Кн. – 2017. – Т. 2. – С. 165-190.
2. Gazzaniga, Michael S, Richard B Ivry, and G. R Mangun. Cognitive Neuroscience: the Biology of the Mind. Fourth edition. New York: W. W. Norton & Company, Inc., 2014.
3. Roux F. E. et al. Language functional magnetic resonance imaging in preoperative assessment of language areas: correlation with direct cortical stimulation //Neurosurgery. – 2003. – Т. 52. – №. 6. – С. 1335-1347.
4. Broca P. Remarks on the seat of the faculty of articulated language, following an observation of aphemia (loss of speech) //Bulletin de la Société Anatomique. – 1861. – Т. 6. – С. 330-57.
5. Eling P. Broca on the relation between handedness and cerebral speech dominance //Brain and language. – 1984. – Т. 22. – №. 1. – С. 158-159.
6. Geschwind N. Carl Wernicke, the Breslau School and the history of aphasia //Selected papers on language and the brain. – Springer Netherlands, 1974. – С. 42-61.
7. Luria A. R. Traumatic aphasia: Its syndromes, psychology and treatment. – Walter de Gruyter, 1970. – Т. 5.
8. Kutas M. et al. Reading senseless sentences: Brain potentials reflect semantic incongruity //Science. – 1980. – Т. 207. – №. 4427. – С. 203-205.
9. Van Berkum J. J. A., Hagoort P., Brown C. M. Semantic integration in sentences and discourse: Evidence from the N400 //Journal of cognitive neuroscience. – 1999. – Т. 11. – №. 6. – С. 657-671.
10. Friederici A. D., Weissenborn J. Mapping sentence form onto meaning: The syntax–semantic interface //Brain research. – 2007. – Т. 1146. – С. 50-58.
11. Osterhout L., Holcomb P. J. Event-related brain potentials elicited by syntactic anomaly //Journal of memory and language. – 1992. – Т. 31. – №. 6. – С. 785-806.
12. Penfield W., Roberts L. Speech and brain mechanisms. – Princeton University Press, 2014.
13. Sidtis D. V. L. et al. Voice and fluency changes as a function of speech task and deep brain stimulation //Journal of speech, language, and hearing research. – 2010. – Т. 53. – №. 5. – С. 1167-1177.
14. Jennum P. et al. Speech localization using repetitive transcranial magnetic stimulation //Neurology. – 1994. – Т. 44. – №. 2. – С. 269-269.
15. Kertesz A. Wernicke–Geschwind model //Encyclopedia of Cognitive Science. – 2001.
16. Bogen J. E., Bogen G. M. Wernicke's region–where is it? //Annals of the New York Academy of Sciences. – 1976. – Т. 280. – №. 1. – С. 834-843.
17. Dronkers N. F. A new brain region for coordinating speech articulation //Nature. – 1996. – Т. 384. – №. 6605. – С. 159.
18. Poeppel D., Hickok G. Towards a new functional anatomy of language //Cognition. – 2004. – Т. 92. – №. 1. – С. 1-12.
19. Chang E. F., Raygor K. P., Berger M. S. Contemporary model of language organization: an overview for neurosurgeons //Journal of neurosurgery. – 2015. – Т. 122. – №. 2. – С. 250-261.
20. Goodale M. A., Milner A. D. Separate visual pathways for perception and action //Trends in neurosciences. – 1992. – Т. 15. – №. 1. – С. 20-25.
21. Hickok G., Poeppel D. The cortical organization of speech processing //Nature reviews. Neuroscience. – 2007. – Т. 8. – №. 5. – С. 393.
22. Rauschecker J. P., Scott S. K. Maps and streams in the auditory cortex: nonhuman primates illuminate human speech processing //Nature neuroscience. – 2009. – Т. 12. – №. 6. – С. 718-724.
23. Makris N. et al. Segmentation of subcomponents within the superior longitudinal fascicle in humans: a quantitative, in vivo, DT-MRI study //Cerebral cortex. – 2004. – Т. 15. – №. 6. – С. 854-869.
24. Maldonado I. L. et al. Surgery for gliomas involving the left inferior parietal lobule: new insights into the functional anatomy provided by stimulation mapping in awake patients //Journal of neurosurgery. – 2011. – Т. 115. – №. 4. – С. 770-779.
25. Saur D. et al. Combining functional and anatomical connectivity reveals brain networks for auditory language comprehension //Neuroimage. – 2010. – Т. 49. – №. 4. – С. 3187-3197.
26. Galantucci S. et al. White matter damage in primary progressive aphasias: a diffusion tensor tractography study //Brain. – 2011. – Т. 134. – №. 10. – С. 3011-3029.
27. Duffau H. et al. Intraoperative mapping of the subcortical language pathways using direct stimulations //Brain. – 2002. – Т. 125. – С. 1-16.
28. Fridriksson J. et al. Impaired speech repetition and left parietal lobe damage //Journal of Neuroscience. – 2010. – Т. 30. – №. 33. – С. 11057-11061.
29. Frey S. et al. Dissociating the human language pathways with high angular resolution diffusion fiber tractography //Journal of Neuroscience. – 2008. – Т. 28. – №. 45. – С. 11435-11444.
30. de Schotten M. T. et al. Monkey to human comparative anatomy of the frontal lobe association tracts //Cortex. – 2012. – Т. 48. – №. 1. – С. 82-96.
31. Leclercq D. et al. Comparison of diffusion tensor imaging tractography of language tracts and intraoperative subcortical stimulations //Journal of neurosurgery. – 2010. – Т. 112. – №. 3. – С. 503-511.
32. Catani M. et al. Perisylvian language networks of the human brain //Annals of neurology. – 2005. – Т. 57. – №. 1. – С. 8-16.
33. Sarubbo S. et al. Frontal terminations for the inferior fronto-occipital fascicle: anatomical dissection, DTI study and functional considerations on a multi-component bundle //Brain Structure and Function. – 2013. – Т. 218. – №. 1. – С. 21-37.
34. Mandonnet E. et al. Does the left inferior longitudinal fasciculus play a role in language? A brain stimulation study //Brain. – 2007. – Т. 130. – №. 3. – С. 623-629.
35. Papagno C. et al. What is the role of the uncinate fasciculus? Surgical removal and proper name retrieval //Brain. – 2010. – Т. 134. – №. 2. – С. 405-414.
36. Duffau H. et al. Is the left uncinate fasciculus essential for language? //Journal of neurology. – 2009. – Т. 256. – №. 3. – С. 382-389.
37. Friederici A. D. et al. The brain differentiates human and non-human grammars: functional localization and structural connectivity //Proceedings of the National Academy of Sciences of the United States of America. – 2006. – Т. 103. – №. 7. – С. 2458-2463.
38. NAESER M. A. et al. Severe nonfluency in aphasia: Role of the medial subcallosal fasciculus and other white matter pathways in recovery of spontaneous speech //Brain. – 1989. – Т. 112. – №. 1. – С. 1-38.
39. Duffau H. et al. Intraoperative subcortical stimulation mapping of language pathways in a consecutive series of 115 patients with Grade II glioma in the left dominant hemisphere. – 2008.
40. Catani M. et al. Short frontal lobe connections of the human brain //cortex. – 2012. – Т. 48. – №. 2. – С. 273-291.
41. Kemerdere R. et al. Role of the left frontal aslant tract in stuttering: a brain stimulation and tractographic study //Journal of neurology. – 2016. – Т. 263. – №. 1. – С. 157-167.
42. Игнатова Ю. П. и др. Современные аспекты изучения функциональной межполушарной асимметрии мозга (обзор литературы) //Экология человека. – 2016. – №. 9.
43. Van der Haegen L. et al. Speech dominance is a better predictor of functional brain asymmetry than handedness: A combined fMRI word generation and behavioral dichotic listening study //Neuropsychologia. – 2013. – Т. 51. – №. 1. – С. 91-97.
44. Gazzaniga M. S., Bogen J. E., Sperry R. W. Some functional effects of sectioning the cerebral commissures in man //Proceedings of the National Academy of Sciences. – 1962. – Т. 48. – №. 10. – С. 1765-1769.
45. Sperry R. Some effects of disconnecting the cerebral hemispheres //Bioscience reports. – 1982. – Т. 2. – №. 5. – С. 265-276.
46. van der Lely H. K. J., Pinker S. The biological basis of language: Insight from developmental grammatical impairments //Trends in Cognitive Sciences. – 2014. – Т. 18. – №. 11. – С. 586-595.
47. Specht K. Mapping a lateralization gradient within the ventral stream for auditory speech perception //Frontiers in human neuroscience. – 2013. – Т. 7.
48. Bemis D. K., Pylkkänen L. Basic linguistic composition recruits the left anterior temporal lobe and left angular gyrus during both listening and reading //Cerebral Cortex. – 2012. – Т. 23. – №. 8. – С. 1859-1873.
49. Friederici A. D. et al. The brain differentiates human and non-human grammars: functional localization and structural connectivity //Proceedings of the National Academy of Sciences of the United States of America. – 2006. – Т. 103. – №. 7. – С. 2458-2463.
50. Price C. J. The anatomy of language: a review of 100 fMRI studies published in 2009 //Annals of the New York Academy of Sciences. – 2010. – Т. 1191. – №. 1. – С. 62-88.
51. Häberling I. S., Steinemann A., Corballis M. C. Cerebral asymmetry for language: comparing production with comprehension //Neuropsychologia. – 2016. – Т. 80. – С. 17-23.
52. Скоромец А.А., Скоромец А.П., Скоромец Т.А. Топическая диагностика заболеваний нервной системы. Руководство для врачей. –СПб.: Политехника, 2014.-628с
53. Miller G. A. et al. Issues in localization of brain function: the case of lateralized frontal cortex in cognition, emotion, and psychopathology //Frontiers in integrative neuroscience. – 2013. – Т. 7.
54. Witteman J. et al. The nature of hemispheric specialization for prosody perception: ERP evidence //Towards a Cognitive Neuroscience of Prosody Perception. – 2014. – С. 63.
55. Van Lancker D. et al. The identification of affective-prosodic stimuli by left-and right-hemisphere-damaged subjects: all errors are not created equal //Journal of Speech, Language, and Hearing Research. – 1992. – Т. 35. – №. 5. – С. 963-970.
56. Schirmer A., Kotz S. A. Beyond the right hemisphere: brain mechanisms mediating vocal emotional processing //Trends in cognitive sciences. – 2006. – Т. 10. – №. 1. – С. 24-30.
57. Mitchell R. L. C., Ross E. D. Attitudinal prosody: What we know and directions for future study //Neuroscience & Biobehavioral Reviews. – 2013. – Т. 37. – №. 3. – С. 471-479.
58. Frühholz S., Gschwind M., Grandjean D. Bilateral dorsal and ventral fiber pathways for the processing of affective prosody identified by probabilistic fiber tracking //Neuroimage. – 2015. – Т. 109. – С. 27-34.
59. Sammler D. et al. Dorsal and ventral pathways for prosody //Current Biology. – 2015. – Т. 25. – №. 23. – С. 3079-3085.
60. Pulvermüller F., Fadiga L. Active perception: sensorimotor circuits as a cortical basis for language //Nature Reviews Neuroscience. – 2010. – Т. 11. – №. 5. – С. 351-360.
61. Möttönen R., Watkins K. E. Motor representations of articulators contribute to categorical perception of speech sounds //Journal of Neuroscience. – 2009. – Т. 29. – №. 31. – С. 9819-9825.
62. D’Ausilio A. et al. Vocal pitch discrimination in the motor system //Brain and language. – 2011. – Т. 118. – №. 1. – С. 9-14.
63. Banissy M. J. et al. Suppressing sensorimotor activity modulates the discrimination of auditory emotions but not speaker identity //Journal of Neuroscience. – 2010. – Т. 30. – №. 41. – С. 13552-13557.
64. Petkov C. I., Logothetis N. K., Obleser J. Where are the human speech and voice regions, and do other animals have anything like them? //The Neuroscientist. – 2009. – Т. 15. – №. 5. – С. 419-429.
65. Hausen M. et al. Music and speech prosody: a common rhythm //Frontiers in psychology. – 2013. – Т. 4.
66. https://4brain.ru/pismo/slovarnyj-zapas.php
67. http://www.mocatest.org/wp-content/uploads/2015/tests-instructions/MoCA-Instructions-Russian_2010.pdf
68. De Deyne S., Verheyen S., Storms G. Structure and organization of the mental lexicon: A network approach derived from syntactic dependency relations and word associations //Towards a theoretical framework for analyzing complex linguistic networks. – 2016. – С. 47-79
69. Collins A. M., Loftus E. F. A spreading-activation theory of semantic processing //Psychological review. – 1975. – Т. 82. – №. 6. – С. 407.
70. Печенкова Е., Фаликман М. Специальная тема выпуска: прайминг и установка //The Russian Journal of Cognitive Science. – 2017. – Т. 4. – №. 1. – С. 4.
71. Vitevitch M. S., Goldstein R. Keywords in the mental lexicon //Journal of memory and language. – 2014. – Т. 73. – С. 131-147.
72. Gahl S., Strand J. F. Many neighborhoods: Phonological and perceptual neighborhood density in lexical production and perception //Journal of Memory and Language. – 2016. – Т. 89. – С. 162-178.
73. Levelt W. J. M. Spoken word production: A theory of lexical access //Proceedings of the National Academy of Sciences. – 2001. – Т. 98. – №. 23. – С. 13464-13471.
74. Levelt W. J. M., Roelofs A., Meyer A. S. A theory of lexical access in speech production //Behavioral and brain sciences. – 1999. – Т. 22. – №. 1. – С. 1-38.
75. Warrington E. K., Shallice T. Category specific semantic impairments //Brain. – 1984. – Т. 107. – №. 3. – С. 829-853.
76. Caramazza A., Shelton J. R. Domain-specific knowledge systems in the brain: The animate-inanimate distinction //Journal of cognitive neuroscience. – 1998. – Т. 10. – №. 1. – С. 1-34.
77. Martin A. et al. Neural correlates of category-specific knowledge //Nature. – 1996. – Т. 379. – №. 6566. – С. 649-652.
78. Taylor K. I., Devereux B. J., Tyler L. K. Conceptual structure: Towards an integrated neurocognitive account //Language and cognitive processes. – 2011. – Т. 26. – №. 9. – С. 1368-1401.
79. Нейман Л.В., Богомильский М.Р. Анатомия, физиология и патология органов слуха и речи: Учеб. для студ. высш. пед. учеб. заведений / Под ред. В.И. Селиверстова. -- М.: ВЛАДОС, 2001. - 224 с.
80. Фундаментальная и клиническая физиология. Учебник. Под ред. А.Г. Камкина, А.А. Каменского. М.: «Академия», 2004.
81. Maclarnon A., Hewitt G. Increased breathing control: another factor in the evolution of human language //Evolutionary Anthropology: Issues, News, and Reviews. – 2004. – Т. 13. – №. 5. – С. 181-197.
82. Winkworth A. L. et al. Breathing patterns during spontaneous speech //Journal of Speech, Language, and Hearing Research. – 1995. – Т. 38. – №. 1. – С. 124-144.
83. Henderson A., Goldman-Eisler F., Skarbek A. Temporal patterns of cognitive activity and breath control in speech //Language and Speech. – 1965. – Т. 8. – №. 4. – С. 236-242.
84. Smith A., Goffman L., Stark R. E. Speech motor development //Seminars in Speech and Language. – © 1995 by Thieme Medical Publishers, Inc., 1995. – Т. 16. – №. 02. – С. 87-99.
85. Barlow S. M., Estep M. Central pattern generation and the motor infrastructure for suck, respiration, and speech //Journal of communication disorders. – 2006. – Т. 39. – №. 5. – С. 366-380.
86. Arbib M. A. 31. Gesture as precursor to speech in evolution//Body-Language-Communication. – 2013. – Т. 1.
87. Hillis A. E., Caramazza A. The graphemic buffer and attentional mechanisms //Brain and language. – 1989. – Т. 36. – №. 2. – С. 208-235.
88. Planton S. et al. The «handwriting brain»: a meta-analysis of neuroimaging studies of motor versus orthographic processes //Cortex. – 2013. – Т. 49. – №. 10. – С. 2772-2787.
89. Binder J. R. et al. Human temporal lobe activation by speech and nonspeech sounds //Cerebral cortex. – 2000. – Т. 10. – №. 5. – С. 512-528.
90. DeWitt I., Rauschecker J. P. Phoneme and word recognition in the auditory ventral stream //Proceedings of the National Academy of Sciences. – 2012. – Т. 109. – №. 8. – С. E505-E514.
91. MacGregor L. J. et al. Ultra-rapid access to words in the brain //Nature Communications. – 2012. – Т. 3. – С. 711.
92. Petersen S. E., Fiez J. A. The processing of single words studied with positron emission tomography //Annual review of neuroscience. – 1993. – Т. 16. – №. 1. – С. 509-530.
93. Nobre A. C. et al. Word recognition in the human inferior temporal lobe //Nature. – 1994. – Т. 372. – №. 6503. – С. 260-263.
94. Puce A. et al. Differential sensitivity of human visual cortex to faces, letterstrings, and textures: a functional magnetic resonance imaging study //Journal of neuroscience. – 1996. – Т. 16. – №. 16. – С. 5205-5215.
95. Dehaene S. et al. The neural code for written words: a proposal //Trends in cognitive sciences. – 2005. – Т. 9. – №. 7. – С. 335-341.
96. Dehaene S. et al. The visual word form area: a prelexical representation of visual words in the fusiform gyrus //Neuroreport. – 2002. – Т. 13. – №. 3. – С. 321-325.
