
Глутаматные рецепторы

Глутамат натрия владеет умами
Как много сплелось в этом слове для нашего человека! Одни боятся «страшного и ядовитого глутамата натрия» как огня, другие же являются ценителями вкуса “умами”. Автор даже слышал истории о том, как некоторые счастливчики смогли выкрасть с работы в колбасном цеху целый килограмм глутамата!
Глутамат (он же глутаминовая кислота) впервые был открыт немецким химиком Карлом Генрихом Раттгаузеном в 1866 году. Он изучал продукты взаимодействия глютена (смеси белков, получаемых из злаковых растений) с серной кислотой. В 1907 году японец Икэда Кикунаэ, изучая состав водорослей комбу и их применение в пищевой промышленности, решил попробовать на вкус кристаллы, оставшиеся после выпаривания бульона из водорослей. К его удивлению, вкус оказался таким же, как и у цельных кусков комбу, используемых в кулинарии. Позднее, идентифицировав состав осадка, Икэда понял, что выделил глутамат. Позднее он улучшил и запатентовал свой метод получения глутамата из водорослей (именно тогда при очистке сырья глутамат начали переводить в форму натриевой соли). Этот метод интересен также и тем, что был применён в первом производстве бульонных кубиков в Японии. Японцы, кстати говоря, очень высоко оценили такой вкус и даже придумали для него слово “умами”, дословно – приятный вкус.
Впервые глутамат был обнаружен в организме животных в 1940-х годах. В 50-х было установлено, что наибольшие его концентрации отмечаются в органах, составляющих нервную систему. В конце 50-х годов группа учёных под руководством Дэвида Кёртиса и Джефри Уоткинса установила, что глутамат может выполнять роль нейротрансмиттера в нервной системе. В своих опытах на изолированном спинном мозге жабы учёные впервые наблюдали эффекты глутамата in vivo: в концентрациях 10-2 – 10-4 М вещество сначала возбуждало нейроны, а затем, подобно открытой и описанной ранее ГАМК, проявляло тормозящие свойства. В дальнейшем те же учёные провели опыт на изолированных клетках Реншоу – вставочных нейронах спинного мозга, регулирующих мышечный тонус путём выброса глицина, тормозного нейромедиатора. Эти клетки к тому времени были достаточно хорошо изучены: о них уже было известно то, что они являются тормозными нейронами и что их активность избирательно стимулируется ацетилхолином. В процессе опытов in vitro выяснилось, что они, как оказалось, могут также быть активированы и глутаматом в достаточных концентрациях! Интересно, что была проведена серия опытов не только с L-глутаматом, который и присутствует в нервной системе в нормальных условиях, но и с его D-изомером. Также были опыты с L- и D-изомерами аспартата, который структурно схож с глутаматом. Таким образом, изначально глутамат и близкие к нему по структуре соединения получили название «возбуждающие аминокислоты» (excitatory amino acids). В 70-х годах было выдвинуто предположение о разделении рецепторов возбуждающих аминокислот на NMDA и не-NMDA. Данное предположение было основано на опытах с единичными нейронами из разных отделов нервной системы: N-метил-D-аспартат в тех же концентрациях, что и глутамат, возбуждал нейроны спинного мозга сильнее, нежели чем нейроны мозга головного. Также выяснилась интересная вещь: избирательные блокаторы NMDA-рецепторов блокировали только их, в то же время неизбирательные блокаторы рецепторов возбуждающих аминокислот подавляли активность нейронов как в спинном, так и в головном мозге. При скрининге активности производных различных аминокислот среди NMDA-рецепторов были выделены ещё два класса – каинатные рецепторы (возбуждаются каиновой кислотой) и АМРА-рецепторы (возбуждаются альфа-амино-3-гидрокси-5-метил-4-изоксазолпропионовой кислотой). В начале 80-х годов выявлена роль глутаматных рецепторов в феномене долговременной потенциации (LTP), когда при повторяющемся выбросе нейромедиаторов с пресинаптического нейрона на постсинаптический он начинает возбуждаться при меньших концентрациях нейромедиатора. Этот эффект лежит в основе явления синаптической пластичности и может быть описан правилом Хэбба: «Если аксон клетки А расположен достаточно близко к клетке Б, чтобы возбуждать её, и постоянно участвует в её активации, то в одной или обеих клетках происходят такие метаболические изменения или процессы роста, что эффективность А как одной из клеток, активирующих Б, повышается». Таким образом, глутаматергические нейроны являются одним из главных звеньев процессов обучения и запоминания. В 90-х, благодаря применению молекулярно-биологических методов изучения работы рецепторов глутамата, они были разделены на ионотропные (iGlu) и метаботропные (mGlu). К ионотропным (т.е. представляющим собой ионный канал) рецепторам отнесли NMDA, AMPA и каинатные. К метаботропным – все остальные, которые разделили на 3 группы в зависимости от наблюдаемых при возбуждении эффектов.
NMDA-рецепторы
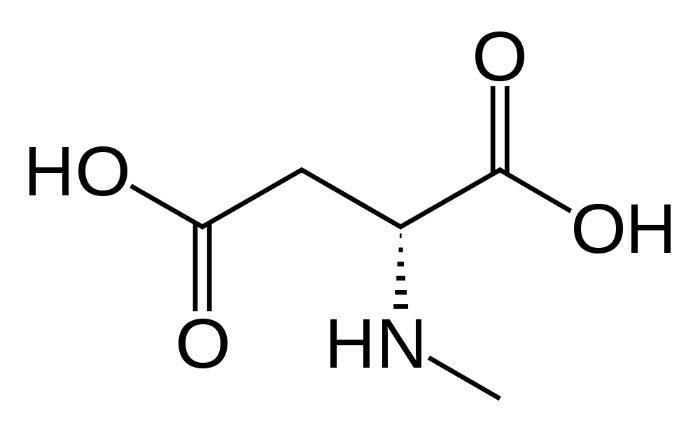
N-метил-D-аспартат
Данный тип рецепторов глутамата представляет собой одновременно потенциал-зависимый и лиганд-зависимый ионный канал, избирательно пропускающий положительно заряженные ионы. Исключение составляют ионы магния и цинка, которые, находясь вне клетки, выступают в качестве блокаторов. Таким образом, основную часть ионного тока составляют ионы кальция и натрия – их данный ионный канал пропускает внутрь клетки, и ионы калия – их он из клетки выпускает. Стоит отметить интересную особенность NMDA-рецептора: он не может быть активирован лишь одной молекулой лиганда. Для открытия ионного канала необходимо два лиганда на разных субъединицах. Лигандами могут быть только D-изомеры аминокислот-нейромедиаторов.
Структурно NMDA-рецептор состоит из четырёх субъединиц, две из которых являются представителями класса NR1, и две другие – представителями класса NR2. В середине нулевых также был выделен новый класс субъединиц – NR3, о нём мы поговорим отдельно.
На внеклеточной части рецептора расположены сайты связывания различных модуляторов и агонистов. Здесь у субъединиц NR2 расположен сайт связывания глутамата, аспартата и собственно N-метил-D-аспартата, а у субъединиц NR1 – сайт связывания ко-агониста – глицина или серина.
Трансмембранная часть формирует ионный канал, и также стоит отметить, что именно эта часть рецепторного белка богата сайтами связывания различных агонистов – от всем известных анестетика кетамина и диссоциатива PCP (фенциклидин) до модельных NMDA-блокаторов, таких как MK810, который часто используют в фармакологических исследованиях. Наконец, внутриклеточная часть NMDA-рецептора: она является регуляторной и может взаимодействовать с различными киназами и фосфатазами, которые путём фосфорилирования или дефосфорилирования влияют на активность ионного канала.
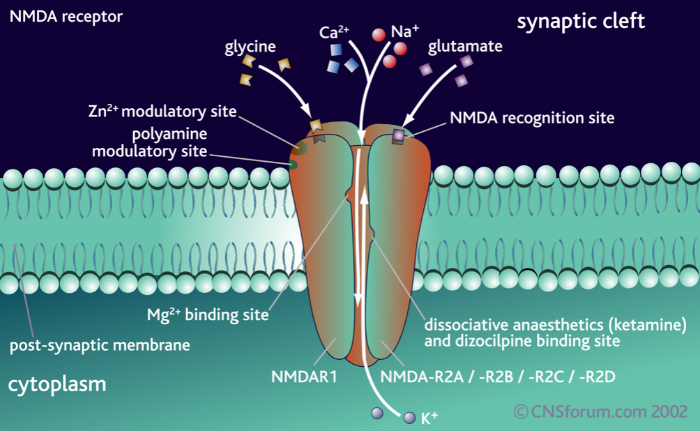
Структура NMDA-рецептора
GluN1, или же NR1 – это название группы белков, составляющих одноимённую субъединицу NMDA-рецептора. Существует в восьми вариантах: NR1-1a, NR1-1b, NR1-2a, NR1-2b, NR1-3a, NR1-3b, NR1-4a, NR1-4b. Из этих субъединиц NR1-1a является самой распространённой. Необходимо отметить также, что все эти варианты субъединиц кодируются одним геном – GRIN1. Такое разнообразие возможно благодаря альтернативному сплайсингу, который в разных частях и клетках нервной системы происходит по-разному. Интересно, что мутации гена GRIN1 ассоциированы с риском развития ранней эпилепсии и риском возникновения острого психоза при приёме метамфетамина и иных психостимуляторов. GluN2, или же NR2 — название группы белков, составляющих другую субъединицу NMDA-рецептора. В отличие от NR1, в ней присутствуют 4 различных белка, каждый из которых кодируется индивидуально отдельным геном: GRIN2A, GRIN2B, GRIN2C, GRIN2D.
С этим типом субъединиц связан интересный феномен. В мозге новорождённых и молодых млекопитающих в составе NMDA-рецепторов преобладает NR2B-субъединица. При активации рецепторов, которые содержат эту субъединицу, соответствующие каналы остаются в открытом положении гораздо дольше. Кроме того, нейроны с такими рецепторами быстрее развивали долговременную потенциацию. Однако с возрастом большая часть NR2B-субъединиц в рецепторных комплексах заменяется на NR2C и NR2A. Это отчасти объясняет снижение с возрастом способностей к обучению, так как долговременная потенциация является одним из ключевых механизмов в процессе запоминания. Не стоит думать, что такая «замена» направлена исключительно во вред организму – в опытах in vivo и in vitro установлено, что клетки с NR2B-субъединицами не имеют никакой защиты от эксайтотоксичности, в то время как преобладание рецепторной субъединицы NR2A значительно повышало порог, при котором наступала клеточная гибель.
GluN3, он же NR3 – это относительно недавно открытая группа белков, существующих в вариантах NR3A и NR3B и являющихся субъединицами NMDA-рецептора. У генов белков субъединиц третьей группы наблюдается наименьшая гомология с остальными генами субъединиц не только NMDA-рецептора, но и других глутаматных ионотропных рецепторов. Интересно также, что гомология между генами белков этой группы меньше, чем гомология внутри остальных групп. Однако, как и во второй группе, соотношение различных вариантов субъединиц также изменяется с возрастом.

Относительное изменение количества различных вариантов NR3-субъединицы с возрастом.
Как видно из вышеприведенных абзацев, NMDA-рецепторы со временем изменяют свой состав и комбинации субъединиц. Это нормальное явление, наблюдаемое не только у человека, но и у других позвоночных. К сожалению, конкретный механизм такой замены ещё не до конца ясен, а самое главное – пока что не выявлены эпигенетические механизмы, лежащие в основе таких изменений.
Лиганды NMDA-рецепторов
В клинике в основном применяются антагонисты NMDA-рецепторов. Самое широкое применение данный фармакологический класс соединений нашёл в неврологии и анестезиологии. Например, стоит отметить такие широко известные соединения, как кетамин, фенциклидин, декстрометорфан. Изначально они использовались для анестезии (кетамин, фенциклидин – под маркой «Сернил») и подавления кашля. Однако их свойства вызывать в немедицинских дозировках интересные нарушения психики были тепло оценены психоделической контркультурной тусовкой, что, собственно, и послужило главной причиной введения контроля за их оборотом. Интересно также, что фенциклидин, он же ПЦП/PCP, рассматривался американскими военными в качестве химического оружия-психотомиметика, способного гуманно снизить уровень боеспособности войск противника. У военных РСР даже получил своё кодовое обозначение – SN, от торгового имени «Сернил» под которым он первое время использовался в клинике.
Помимо этих известных препаратов, в клиническом использовании до сих пор находятся флупиртин (анальгетик) и мемантин (используется для лечения деменций различного генеза). Интересно также, что в опытах на животных NMDA-антагонисты эффективно предотвращали гибель нейронов от эксайтотоксичности, однако при длительном и частом введении в больших количествах они вызывали специфическое поражение ЦНС, названное лезиями Олни (в честь учёного, впервые их описавшего – Джона Олни). В отдельных публикациях были описаны т.н. пограничные морфологические состояния. Например, у крыс после длительной анестезии оксидом азота (I) наблюдалась вакуолизация цитоплазмы нейронов – обычно после данного процесса происходит гибель нейронов путем некроза или апоптоза.
Агонисты NMDA-рецепторов в клинике практически не используются. В 2015 году был зарегистрирован препарат небогламин. В опытах на животных он показал способность ускорять обучение и процессы запоминания. Также в процессе испытаний была выявлена способность небогламина избавлять от кокаиновой зависимости.
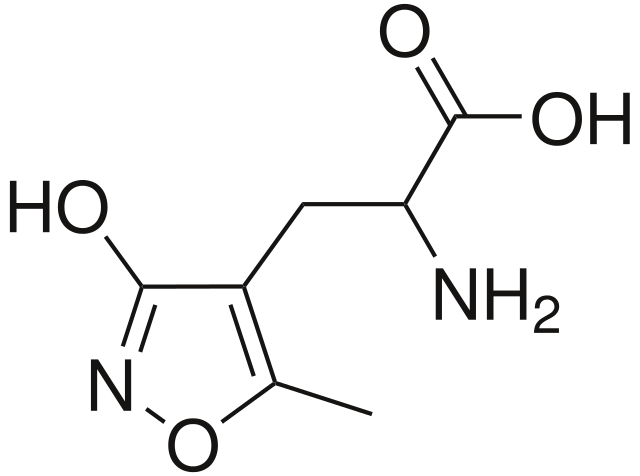
Этот подтип ионотропных глутаматных рецепторов, способных пропускать положительно заряженные ионы, был назван AMPA в честь избирательного агониста - альфа-амино-3-гидрокси-5-метил-4-изоксазолпропионовой кислоты. Как и NMDA, AMPA-рецептор является гетеромерным тетрамером, т.е. состоит их четырёх различных субъединиц: GluR1, GluR2, GluR3, и GluR4. Каждая из субъединиц кодируется своим собственным геном. Стоит отметить, что зачастую AMPA, как и NMDA-рецепторы, состоят не из всех четырёх разных субъединиц, а из сочетания двух одного типа, и двух – другого.
Рецепторы, содержащие в своём составе GluR1-субъединицу, обнаруживаются чаще всего в гранулярных и пирамидальных клетках гиппокампа.
Субъединица GluR2 является необходимой для группы АМРА-рецепторов, которые не пропускают ионы кальция. Интересно, что перед началом трансляции мРНК гена данной субъединицы взаимодействует с белком ADAR (аденозилдезаминаза РНК), который заменяет триплет CAG на CIG (аденозин заменяется на инозин). Соответствующие изменения наступают и в структуре белка – глутамин в 607-й позиции заменяется на аргинин, также, в 764 позиции аргинин заменяется на глицин. В опытах на трансгенных мышах, у которых данный процесс редактирования мРНК был нарушен и АМРА-рецепторы с GluR2-субъединицей были проницаемы для ионов кальция, у животных наблюдались симптомы, схожие с амиотрофическим боковым склерозом. Данная замена аминокислот затрагивает структуру самого ионного канала, а точнее – сайта, ответственного за пропуск ионов с зарядом 2+. Таким образом, нейроны с дефектной GluR2-субъединицей, пропускавшей ионы кальция, были подвержены «физиологической» эксайтотоксичности вследствие повышенной концентрации глутамата в синапсе, которая в нормальных условиях не возникала благодаря избирательной блокаде входа кальция в клетку через конкретно этот тип каналов. Интересно, что при полном выключении редактирования мРНК GluR2, когда не происходило замен глутамин-аргинин и аргинин-глицин, трансгенные мыши умирали в младенчестве от судорог. Такая же замена была обнаружена и при изучении синтеза субъединиц каинатных рецепторов GluR5 и GluR6, однако у них данная замена, наоборот, увеличивает пропускную способность ионного канала в отношении Ca2+. Антитела к GluR2 также используются при иммуногистохимическом выявлении солитарной фиброзной опухоли (SFT) и выбухающей дерматофибросаркомы (DFSP).
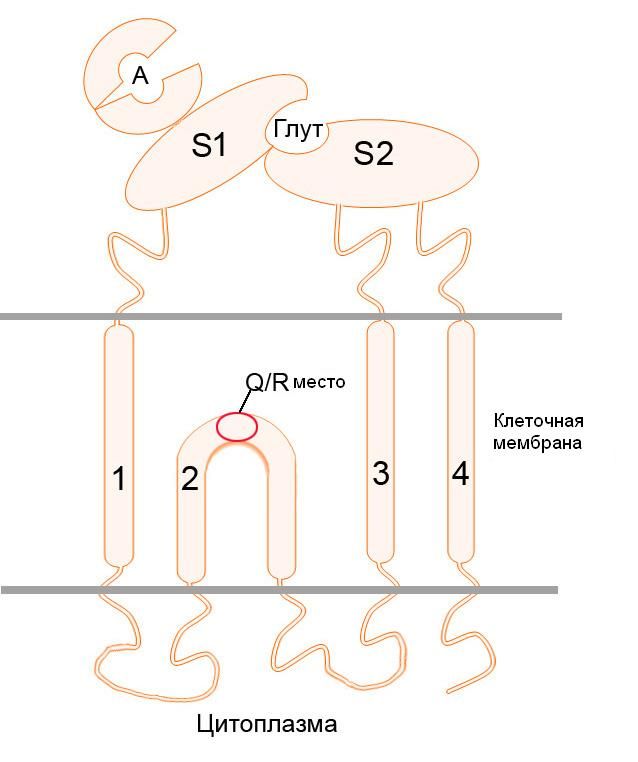
Структура AMPA-рецептора, Q/R – название места замены глутамина на аргинин.
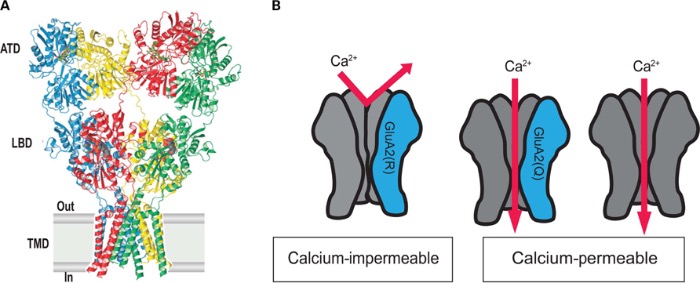
Рецепторы с GluR2, прошедшим на уровне мРНК правильную обработку, непроницаемы для ионов кальция.
GluR3-субъединица.
Эта субъединица, как и GluR2, проходит редактирование на уровне мРНК. Замена в данном случае также происходит благодаря ферментам группы ADAR. В данном случае – ADAR3, который находится исключительно в тканях нервной системы. Однако здесь замена носит необязательный характер – лишь в 80% случаев мРНК субъединиц GluR2 проходят соответствующее редактирование. Разница между отредактированными и неотредактированными субъединицами никак не отражается на выживаемости клетки: у рецепторов с отредактированными субъединицами состояние десенсибилизации длится короче.
GluR4-субъединица.
Специфических отличий от GluR2 и GluR3 субъединиц нет. У GluR4 обнаружена высокая гомология с генами этих субъединиц. Так же, как и GluR3, может встречаться в редактированном и неотредактированном варианте, в обоих случаях влияния на выживаемость нейрона не обнаружено. Согласно исследованиям японских генетиков, у гена субъединицы GluR4 существует 6 SNP-мутаций, которые повышают риск развития шизофрении.
АМРА, наряду с NMDA-рецепторами, является одним из главных звеньев в механизме долговременной потенциации, а значит, и в механизме запоминания. Долговременная потенциация (LTP) выражается в увеличении возбуждающего постсинаптического потенциала в ответ на какой-либо повторяющийся стимул. Согласно современным теориям, LTP в первую очередь связывают с апрегуляцией (увеличением плотности) АМРА-рецепторов на мембране постсинаптического нейрона.
Упрощённо механизм апрегуляции выглядит примерно так: на постсинаптическом нейроне находятся NMDA и АМРА-рецепторы и глутамат возбуждает АМРА-рецептор (NMDA в это время остаётся закрытым благодаря иону магния в ионном канале), и в клетку устремляются ионы натрия, что вызывает деполяризацию. Деполяризация мембраны как бы выталкивает магний из ионного канала в NMDA-рецепторе, таким образом открывая его для кальция. Кальций, входя в клетку, запускает каскад реакций, ведущий к увеличению плотности АМРА-рецепторов на мембране. Кроме того, кальций запускает работу киназы CaMKII, которая путём фосфорилирования регуляторных участков уже существующих АМРА-рецепторов увеличивает их проводимость.
Установлено, что АМРА-рецепторы, наряду с каинатными, являются также и ключевым звеном, задействованным в патогенезе судорожного синдрома. В настоящее время ведутся разработки новых противоэпилептических препаратов, избирательно блокирующих активность АМРА-рецепторов.
Как и NMDA, и АМРА-рецепторы, каинатный рецептор также является ионотропным. Своё название он получил благодаря каиновой кислоте, которая является первым открытым избирательным агонистом рецепторов данной подгруппы.
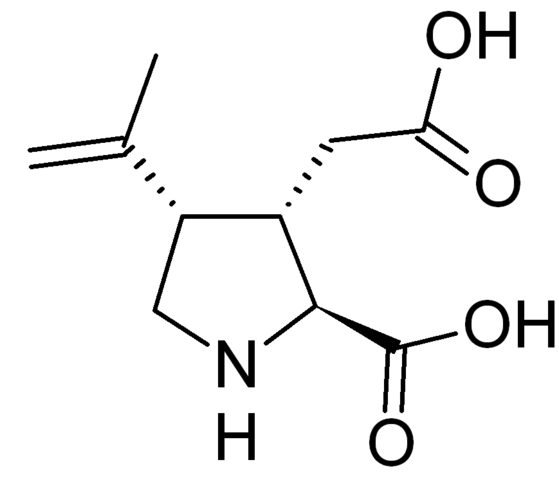
Каиновая кислота
Каинатный рецептор состоит четырёх субъединиц, так же как и NMDA и АМРА. Однако не всё так просто – этих субъединиц на самом деле пять, но они могут быть совмещены в тетрамерных комплексах различного состава и по определённым правилам. Каинатный рецептор формирует ионный канал, проницаемый для натрия и кальция.
Стоит отметить, что после 2009 года произошло переименование субъединиц:
Glu5 теперь GluK1Glu6 теперь GluK2
Glu7 теперь GluK3
KA1 теперь GluK4
KA2 теперь GluK5
Теперь о правилах компоновки – Glu1-3 могут формировать гомотетрамеры, т.е. рецепторы, состоящие, например, только из GluK2 субъединиц. Однако GluK4 и GluK5 такой способности лишены и могут присутствовать лишь в составе гетеротетрамеров (одна GluK4 и три GluK3, например).
GluK1
Эта субъединица каинатного рецептора, которая, как и некоторые субъединицы АМРА-рецепторов, подвергается редакции на уровне мРНК. Интересно, что характер наиболее часто встречающейся замены, а именно – глутамина на аргинин, ведёт к абсолютно тем же последствиям – утрате способности рецептора, содержащего такую субъединицу, пропускать ионы кальция. В отличие от АМРА-рецепторов, отсутствие или наличие отредактированных субъединиц не влияет на выживаемость нейрона. Более того – в отдельных частях нервной системы (спинной мозг, мозолистое тело, мозжечок) редактируется лишь 50% субъединиц, в других же частях (таламус, миндалевидное тело, гиппокамп) редакции подвергается 70%. Кроме того, отредактированные субъединицы могут быть избирательно заблокированы некоторыми жирными кислотами - арахидоновой и докозагексаеновой.
GluK2
Данная субъединица также подвергается редактированию в процессе синтеза. Однако даже тут есть свои уникальные отличия – например то, что процент редактирования непрерывно растёт в процессе эмбрионального развития: начиная от полного отсутствия на стадиях закладывания нервной трубки, заканчивая 80% редакцией всех GluK2-субъединиц обнаруженных в мозге новорождённого. Из этих 80% отредактированных рецепторов 90% приходится на расположенные в сером веществе, остальные 10% - на те, что в белом веществе. Также у этой субъединицы присутствует своя отличительная черта среди всех редактируемых субъединиц – у неё редакция ведёт, наоборот, к увеличению проницаемости ионного канала для кальция. Как уже отмечалось выше, каинатные рецепторы являются одним из ключевых звеньев в возникновении судорожного синдрома. И, как показали исследования, в этом больше задействованы рецепторы, содержащие GluK2-субъединицу – трансгенные мыши, у которых выключена редакция этой субъединицы, показывали сниженный судорожный порог.
GluK3
Эта субъединица изучена чуть меньше, однако уже известны мутации (замена серина в 310 позиции на аланин), которые повышают риск развития белой горячки после запоев и шизофрении. Также описан случай, когда при генетическом обследовании пациента с отставанием в развитии обнаружилась микроделеция 1p34.3 (регион 34.3, малого плеча 1-й хромосомы), включающий в себя и ген этой субъединицы. Так как в указанном регионе находился только один ген, чьё участие в процессах, происходящих в нервной системе, не подвергается сомнению, то причину отставания в развитии связали именно с его отсутствием.
GluK4
Эта субъединица изучена больше в силу исторических причин – она долгое время считалась первой истинно-каинатной. Таким образом, функции этой субъединицы наиболее изучены клиницистами. Например, выявлены мутации гена этой субъединицы, которые влияют на восприимчивость пациента к циталопраму. Как уже упоминалось выше, каинатные рецепторы являются ключевым звеном патогенеза судорог, но в целом на выживаемость нейрона при эксайтотоксичности не влияют. Так вот эта субъединица является исключением – при избыточной активации рецепторов, её содержащих, эксайтотоксичность наступает. Это было подтверждено опытами на мышах (забавный факт – избирательных блокаторов GluK4 на момент проведения опыта в 2009 году не существовало, исследователи пользовались моноклональными антителами к данной субъединице) – при выключении этой субъединицы нейроны дольше выживали в условиях индуцированной эксайтотоксичности.
GluK5
Я, признаться, был несколько удивлён – несмотря на то, что данная субъединица была открыта еще в 1995 году, по ней крайне мало информации. О клиническом значении этой субъединицы существует лишь одна публикация, и то – с отрицательными результатами исследований корреляции различных SNP в её гене с риском психических и неврологических заболеваний. Однако, существует публикация 2013 года, за авторством учёных из Университета Бордо. Они показали, что фосфорилирование GluK5 с помощью уже упомянутой ранее киназы CaMKII регулирует плотность рецепторов на постсинаптической мембране.
Касательно каинатных рецепторов, для того, чтобы ещё раз подчеркнуть их важность и функции, следует упомянуть такой нейротоксин, как домоевая кислота. Это соединение является сильным агонистом ионотропных глутаматных рецепторов, в основном связывается с каинатными рецепторами гиппокампа, вызывая избыточную активацию этих самых рецепторов и, таким образом, оказывая эксайтотоксическое действие. Помимо общей неврологической симптоматики, частым симптомом отравления является временная антероградная амнезия – неспособность запоминать новую информацию. Механизм действия этого токсина прекрасно подтверждает как роль гиппокампа в процессах запоминания, так и роль глутаматных рецепторов в процессе установления долговременной потенциации.
Про ионотропные глутаматные рецепторы, в целом следует также рассказать о «молчащих синапсах». Это «несозревшие» синапсы, которые содержат NMDA-рецепторы, однако практически лишены АМРА-рецепторов. В таких случаях, даже если глутамат и ко-активатор связываются с NMDA-рецептором, он остаётся закрытым, ионный канал для кальция заблокирован ионом магния – который может отсоединиться лишь в случае смены потенциала благодаря работе АМРА-рецепторов, которых в данном случае нет. Именно поэтому такой синапс и называется «молчащим». Такие синапсы постепенно могут перейти в нормальное активное состояние – достаточно вызвать сильную деполяризацию мембраны, чтобы освободить ионный канал в NMDA-рецепторе для кальция, который затем активирует CaMKII и кальмодулин. А они уже через цепь посредников запустят процесс экзоцитоза везикул с AMPA-рецепторами, в результате которого эти рецепторы будут выставлены на постсинаптической мембране, делая синапс полноценным.
И ещё – среди ионотропных рецепторов глутамата выделено два рецептора-сироты – GluD1 и GluD2. Эндогенные лиганды этих рецепторов ещё не найдены, глутаматом они не активируются, а в эту группу отнесены были лишь по доброте душевной и высокой гомологии. С помощью синтетических лигандов даже выяснена их роль – они отвечают за развитие синаптических связей в мозжечке, главным образом находясь в клетках Пуркинье. В опытах на животных мыши с определённым мутантным вариантом этого рецептора умирали вскоре после рождения – при исследовании мозга было обнаружено значительное уменьшение объёма заднего мозга. Люди добрые, не проходите мимо! Помогите этим рецепторам найти свои эндогенные лиганды!
Метаботропные рецепторы разделены на три группы в зависимости от их механизма работы, гомологии структуры и перечня селективных лигандов. Всего существует три группы:
- mGluR1, mGluR5
- mGluR2, mGluR3
- mGluR4, mGluR6, mGluR7, mGluR8
Во-первых, группы рецепторов различаются по механизму действия. Рецепторы первой группы связаны с Gq-белком. При активации тот запускает фермент фосфолипазу С (PLC), который осуществляет расщепление фосфатидилинозитол-4,5-фосфата (PIP2) на диацилглицерол (DAG) и инозитол-1,4,5-трифосфат (IP3). DAG в свою очередь активирует протеинкиназу С (PKC), которая осуществляет фосфорилирование других белков; а IP3 служит агонистом IP3-рецепторов, которые запускают высвобождение кальция из эндоплазматического ретикулума, таким образом, ионы кальция активируют кальмодулин-зависимые протеинкиназы, также участвующие в фосфорилировании белков.
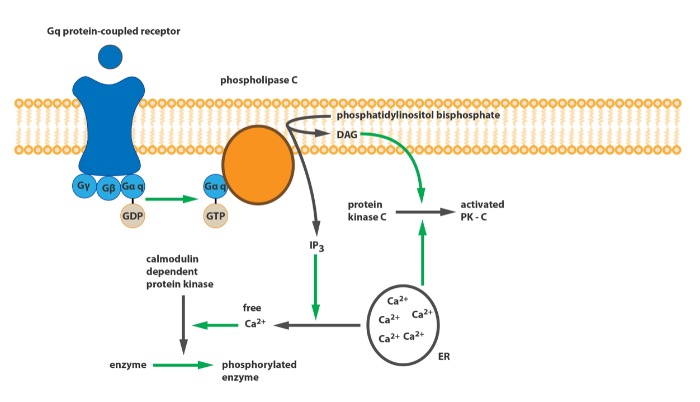
Принципиальная схема работы Gq-связанного рецептора
Остальные группы глутаматных рецепторов, вторая и третья, осуществляют своё действие через Gi-белок. Это означает, что при активации этих рецепторов блокируется работа аденилатциклазы, которая в активном состоянии превращает АТФ в цАМФ. Следовательно, останавливается работа цАМФ-зависимой протеинкиназы (второе название – протеинкиназа А, PKA), и каскад реакций фосфорилирования, изменяющих гомеостаз кальция, не запускается. Исходя из вышеизложенного, можно уяснить различия в эффектах этих рецепторов: при активации рецепторов 1 группы активность NMDA и АМРА-рецепторов увеличивается (в т.ч. за счёт повышения синаптической плотности), но также увеличивается восприимчивость к эксайтотоксичности; при активации рецепторов 2 и 3 групп, наоборот, активность и плотность ионотропных рецепторов уменьшается, также уменьшается вероятность возникновения эксайтотоксического возбуждения.
Можно заключить всё в небольшую таблицу.
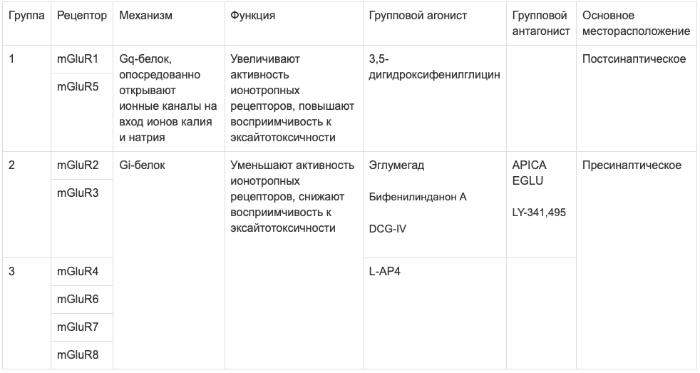
Касательно групповых агонистов стоит сделать ещё небольшую заметку. Агонисты рецепторов 2 группы в основном проявили себя как антипсихотики – например, бифенилинданон А (BINA) даже смог купировать модельный психоз, вызванный ДОБом, DCG-IV показал себя как антиконвульсант. Однако, оба эти вещества нарушали процессы запоминания. Причём, в отличие от домоевой кислоты, чьё амнестическое действие возникает из-за прямой индукции эксайтотоксичности в нейронах гиппокампа, агонисты метаботропных рецепторов 2 группы лишь блокировали «молекулярный субстрат», необходимый для возникновения долговременной потенциации. Таким образом, можно разделить метаботропные рецепторы на возбуждающие и тормозные, хотя такое разделение лишь условно.
Говорить об отдельных рецепторах можно и вне контекста группы, однако отдельные особенности рассмотреть однозначно стоит.
1 группа
mGluR1
Находится на адренергических нейронах и является положительным модулятором активности соответствующих рецепторов – селективные антагонисты mGluR1 оказывают адренолитическое действие. В исследованиях на мышах, лишённых гена данного рецептора, выявлено отсутствие каких-либо макро- и микроскопических патологий. Однако в тестах на координацию движений и запоминание мыши без этого рецептора показали худшие результаты по сравнению с контрольной группой. Интересно, что имеются сообщения о случаях, когда при болезни Ходжкина организм пациента вырабатывал аутоантитела к mGluR1, что выражалось в виде паранеопластической атаксии. Мутации гена этого рецептора также могут быть причиной рецессивной спиноцеребеллярной атаксии.
mGluR5
Этот метаботропный рецептор интересен тем, что является одной из мишеней лития в организме. Литий (в форме кислых органических солей или карбоната) применяется в медицине в качестве нормотимика. На рецепторе имеется три сайта связывания – один для глутамата, а два других – для модуляторов и экзогенных лигандов, что делает mGluR5 перспективной мишенью для разработки нормотимиков, антидепрессантов и транквилизаторов. Например, лиганды этого рецептора, лишённые способности проникать через ГЭБ, исследуются в качестве средств, влияющих на тонус мочевого пузыря. Однако клиническое использование антагонистов и негативных модуляторов mGluR5 скорее всего будет ограниченным, так как уже в исследованиях на крысах данный класс соединений наряду со способностью снимать тревогу показал негативный побочный эффект – нарушения памяти. Агонисты mGluR5 исследуются в качестве средств против деменции и для восстановления после травм, уже опубликованы положительные результаты с 2-хлоро-5-гидроксифенилглицином в качестве перспективного агониста.
2 группа
mGluR2
Этот рецептор является «тормозным», как и большинство тех, что будут описаны ниже. В данном случае антагонисты и негативные модуляторы являются перспективными антидепрессантами - MGS-0039, например. Клиническое значение – в некоторых случаях амиотрофического бокового склероза описана низкая плотность mGluR2-рецепторов на нейронах. Скорее всего, это и явилось одной из причин гибели нервных клеток в случаях с теми пациентами, т.к. данные рецепторы защищают нейроны от гибели, в том числе и от эксайтотоксичности. Также данный рецептор способен взаимодействовать с 5-HT2a-рецептором серотонина. При посмертном исследовании мозга больных шизофрений было открыто, что при некоторых вариантах заболевания увеличение плотности (upregulation) 5-HT2a-рецепторов коррелирует с уменьшенной плотностью (downregulation) mGluR2-рецепторов, которые являются тормозными, в т.ч. и в случае расположения на серотонинэргических нейронах.
mGluR3
Мутации гена этого рецептора повышают риск развития биполярного расстройства, шизофрении и склонности к зависимостям. Соответственно, как и mGluR2, данный рецептор также является мишенью для различных экспериментальных препаратов. Более того, благодаря высокой гомологии (в т.ч. и сайта связывания) в научной литературе некоторые групповые лиганды обозначаются как mGluR2/3-агонисты (или антагонисты). Также мутации в гене этого рецептора могут влиять на риск развития меланомы – в 16,3% случаев меланомы, у пациента обнаруживается мутация mGluR3, выражающаяся в замене глутамина в 870 позиции на лизин.
3 группа
mGluR4
Этот рецептор интересен тем, что помимо ЦНС находится также и… на языке! Да, именно благодаря нему человек способен чувствовать тот самый вкус «умами», благодаря которому и началась история глутамата. Также, помимо обычных для таких рецепторов функций (LTP, участие в процессах запоминания и координации движений), mGluR4 обнаружен и в иммунных клетках – Т-хелперах. При активации этих рецепторов Т-хелпер снижает продукцию интерлейкина-17. Мыши, лишённые этого рецептора, были более восприимчивы к экспериментальным формам энцефаломиелита и рассеянного склероза.
mGluR6
Данный подвид глутаматного рецептора найден в больших количествах в сетчатке. Клетки, имеющие этот рецептор (в большинстве своём – ON-подтип биполярных клеток, у них рецептор располагается постсинаптически), участвуют в регуляции силы восприятия светового раздражителя. Некоторые мутации гена этого рецептора ведут к никталопии (народное название – «куриная слепота»), нарушению сумеречного зрения.
mGluR7
Большая плотность этих рецепторов наблюдается в нейронах слуховых органов и вестибулярного аппарата. Также, благодаря селективному антагонисту - XAP044, mGluR7-рецепторы были обнаружены в миндалевидном теле. Мыши, у которых отсутствовал этот рецептор, ( + NB) и те, у которых он был заблокирован упомянутым веществом, были меньше подвержены стрессу и беспокойству, однако у них была замедлена выработка рефлексов.
mGluR8
Об этом рецепторе известно мало. Однако, исходя из его близкого расположения к региону 7-й хромосомы, который подвергается делеции при синдроме Смита-Лемли-Опица, высказаны предположения о том, что его отсутствие влияет на неврологический статус пациента, в т.ч. обуславливая умственную отсталость, наблюдаемую при этом заболевании.
Как вы могли уже выяснить из прочтённого, рецепторы глутамата играют огромную роль в регуляции функционирования и развития нервной системы. Например, глутамат играет роль в гибели нейронов в условиях гипоксии – в таких условиях транспортёр глутамата (EAAT) просто не способен осуществлять обратный захват нейромедиатора в клетку. Таким образом, при массовой гибели нервных клеток количество высвободившегося глутамата растёт в геометрической прогрессии, вызывая эксайтотоксичное возбуждение в ещё живых нейронах, которые тоже могут умереть из-за этого.
Глутатэргическая система играет роль ключевого звена во множестве неврологических и психиатрических заболеваниях – у более чем 30 нозологических единиц заболеваний доказана роль нарушений метаболизма глутамата в процессе патогенеза. Описать их всех в рамках единичной публикации, увы, невозможно – интересующихся отсылаю к книге «Нейропсихофармакология антагонистов NMDA-рецепторов» (авторы - Беспалов А.Ю., Звартау Э.Э.), эта книга является одной из самых информативных из русскоязычных.
Стоит рассказать об отечественных наркологах Крупицком и Гриненко и их изобретении – кетаминовой терапии алкоголизма. Само название звучит довольно интересно. Это в прямом смысле шоковая терапия – согласно теории авторов, после сильного психоэмоционального переживания, связанного с алкоголем, у пациента пропадёт тяга к алкоголю. Терапия проводится в три этапа:
- С пациентом проводится просветительская беседа о вреде алкоголя, его пагубном влиянии на качество жизни. Также на этом этапе пациенту объясняют, что от введённых препаратов он будет испытывать галлюцинации, связанные с алкоголем, которые изменят его отношение к пьянству.
- Пациенту вводится бемегрид (блокатор ГАМК-рецепторов), этимизол (дыхательный аналептик) и кетамин. По сути, помимо галлюциногенного действия данная смесь является анксиогенной – вызывающей тревогу и страх. А затем пациента нужно немного попугать, цитирую – «Далее на фоне эмоциогенной, драматической по характеру музыки проводят психотерапевтические воздействия на больного, находящегося в состоянии легкого кетаминового наркоза, во время которого определенный раппорт с ним сохраняется». Подбирать угрозы авторы советуют исходя из индивидуального анамнеза и истории болезни. Авторы рекомендуют на пике эмоциональных переживаний дать пациенту понюхать или даже подержать во рту немного алкоголя.
- На следующий день после процедуры с пациентами проводится групповая психотерапия, где они делятся пережитым опытом, пересказывают друг другу свои галлюцинации и обсуждают вынесенный опыт. Довольно интересное изобретение, что-то на уровне атропинокоматозной терапии. Самое интересное, что процедура действительно обладала большой эффективностью и после неё было относительно мало рецидивов.
В качестве заключения могу сказать, что глутаматэргическая система мозга – это одна из самых широкоспециализированных сигнальных систем в нашем мозге и нервной системе, и её роль действительно сложно переоценить.
Источники:
- https://www.ncbi.nlm.nih.gov/pubmed/12467378 Neuroscientist. 2002 Dec;8(6):562-73. GABA and glutamate in the human brain. Petroff OA1.
- http://old.www.ncbi.nlm.nih.gov/pubmed/16402093 Br J Pharmacol. 2006 Jan;147 Suppl 1:S100-8. The glutamate story.
