
Боль: ноцицепция

Когда кто-то говорит “больно”, мне часто вспоминается сцена в начале фильма “Майор Пэйн”. Кадр плавно переходит от ног главного героя наверх, показывая его крупным планом. Повсюду взрывы, крики, пыль, звуки выстрелов. Майор подходит к раненому солдату, который вопит: “Рука! Меня ранили!”. Пэйн садится и спокойно просит дать посмотреть место ранения. “Хочешь покажу, как забыть про боль в ране? Дай руку, немного усилий и все”. С этими словами Майор хладнокровно ломает палец солдату, на что раздается крик: “Палец! Мой Палец!”. Со словами “Проверенное средство!” главный герой поднимается и выходит из кадра.
Действительно, когда боль приходит в нашу жизнь, она занимает в ней первое место. Она контролирует движения и овладевает личностью. Независимо от нашего происхождения, возраста и положения каждый способен рассказать об ощущении боли. Но мало кто задумывался об её истинной природе, механизмах и физиологии. Почему боль может ослабевать и усиливаться, как организм узнаёт о боли, чем она может быть полезна, что такое нейропластичность, а также каким образом боль поддаётся лечению — в следующем ноцицептивном обзоре.
По эпидемиологическим данным острая боль различной интенсивности возникает в течение жизни у 80–90% людей , а хроническая беспокоит от 24% до 46% всех людей, 15% из которых считают ее непереносимой. Источником боли в 90% случаев является опорно-двигательный аппарат. Следует отметить, что боль всегда субъективна и часто требует уникального лечения. Согласно IASP, боль — неприятное сенсорное и эмоциональное переживание, связанное с истинным или потенциальным повреждением ткани или описываемое в терминах такого повреждения.
Классификация
Существует немало вариантов классификации боли, вот несколько из них. По происхождению боль бывает центрального и периферического генеза. По площади и объёму болевого восприятия она может быть местной (локализованной) и системной (общей, разлитой). В зависимости от глубины повреждения тканей различают следующие виды боли:
- поверхностная соматическая — при повреждении кожных покровов и слизистых оболочек;
- глубокая соматическая — при повреждении костей, суставов, скелетных мышц, соединительной ткани;
- глубокая висцеральная — при растяжении, сдавлении, повреждении внутренних органов.
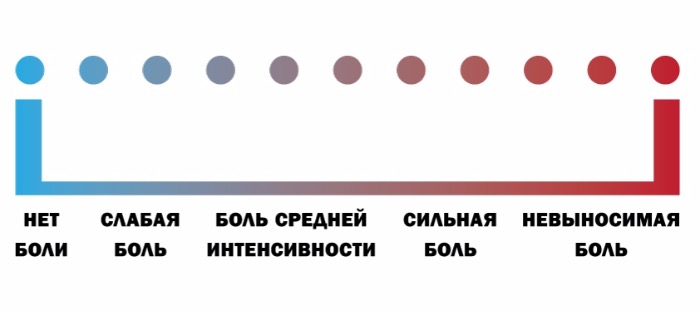
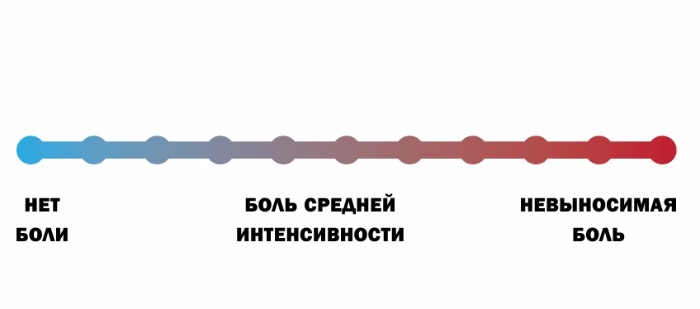
По интенсивности различают слабую, среднюю и сильную боль. По степени переносимости боль может быть легко переносимой (если она слабая), трудно переносимой (если она сильная), непереносимой (нетерпимой). Что касается клиники, для нее большое значение имеет так называемая шкала оценки боли. В зависимости от того, способен ли пациент самостоятельно рассказать о болевых ощущениях, существуют следующие шкалы:
а) если способен — вербальная рейтинговая (ВРШ) (рис Б), цифровая рейтинговая (ЦРШ) (рис. В) и визуальная аналоговая (ВАШ) (рис. Г) шкалы;

При использовании вербальной шкалы у пациента необходимо выяснить, испытывает ли он какую-либо боль прямо сейчас. Если боли нет, то его состояние оценивается в 0 баллов. Если наблюдаются болевые ощущения, необходимо спросить: «Вы могли бы сказать, что боль усилилась, или она невыносимая, или это самая сильная боль, которую вы когда-либо испытывали?» Если это так, то фиксируется самая высокая оценка в 10 баллов (рис. В). Если же нет ни первого, ни второго варианта, то далее необходимо уточнить: «Можете ли вы сказать, что ваша боль слабая, средняя (умеренная, терпимая, несильная), сильная (резкая) или очень (особо, чрезмерно) сильная (острая)» (рис. Б и Г).
б) если не способен — шкала оценки боли по выражению лица (рис. А).
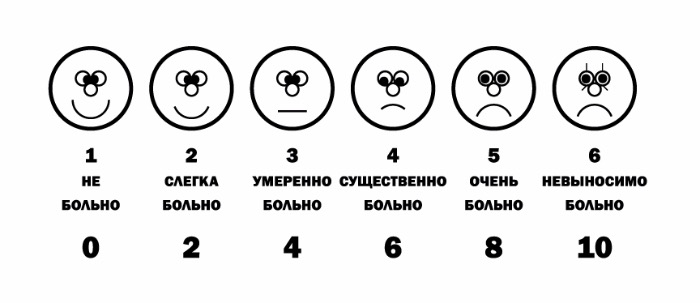 Оценка боли по выражению лица
Оценка боли по выражению лица
 Вербальная рейтинговая шкала
Вербальная рейтинговая шкала
 Цифровая рейтинговая шкала (1-10)
Цифровая рейтинговая шкала (1-10)
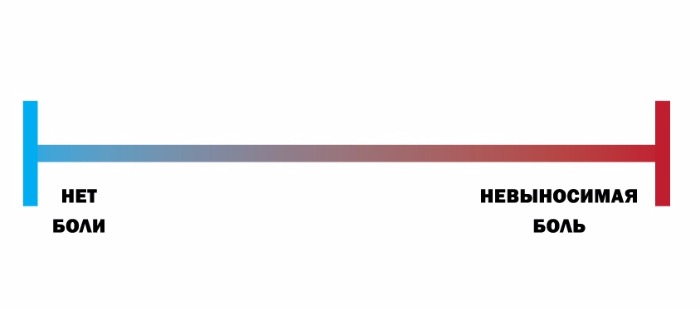 Цифровая аналоговая шкала
Цифровая аналоговая шкала
По характеру, качеству и субъективным ощущениям боль бывает следующих видов:
- приступообразная, пароксизмальная, пульсирующая;
- тупая, колющая, режущая, стреляющая, давящая, сжимающая, рвущая, дёргающая, сверлящая, жгучая (казуалгия);
- проекционная, иррадиирующая, перемежающаяся, отражённая, реактивная, гемиалгия, фантомная и др.
Помимо этого, по некоторым данным , боль делят на острую (меньше 4-х недель), подострую (от 4-х недель до 3-х месяцев) и хроническую (от 3-х до 6-ти месяцев).
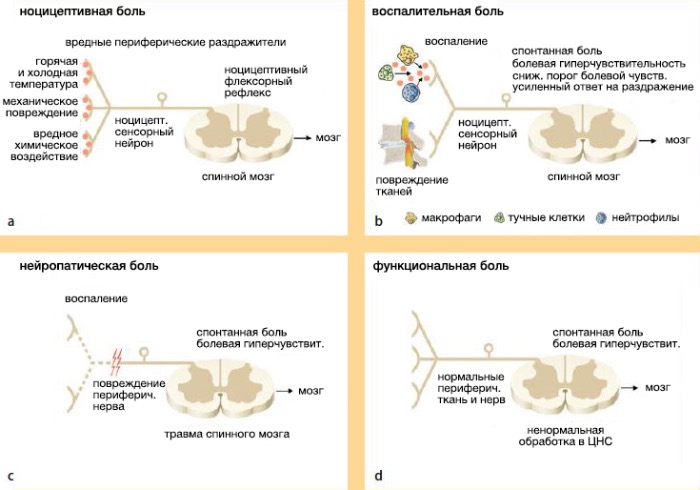
Острая боль вызвана адекватной стимуляцией ноцицептивных нейронов. Обычно возникает при повреждении мягких тканей и сопровождает процесс регенерации тканей . Подострая боль зачастую не настолько интенсивна и следует за острой фазой. Она относится к органической боли, возникающей в процессе заживления тканей и их ремоделирования, и длится не более 12 недель. Хроническая боль, наоборот, утратила свою протекторную роль. У пациентов, страдающих таким видом боли, часто возникают трудности с определением повреждения ткани, или отсутствует стимул, который стал изначальным источником боли. Хроническая боль способствует биохимическим и фенотипическим изменениям (молекулярным и клеточным) в нервной системе, приводящим к физиологической, метаболической и иммунологической альтерации, которая, как известно, угрожает нормальному протеканию процессов гомеостаза, способствует развитию болезни и вследствие этого приводит к смерти . Современная классификация, предложенная Клиффордом Вульфом , предлагает следующую дифференцировку боли: ноцицептивная, воспалительная, невропатическая и функциональная.
Ноцицептивная боль является жизненно важным физиологическим показателем, ощущение которой возникает при травмах . Острая ноцицептивная боль возникает при воздействии на ткань болевого раздражителя, достаточного для повреждения. Это имеет важное значение для защиты ткани от дальнейшего повреждения, так как вызывает определённый рефлекс. Несмотря на основную функцию ноцицептивной системы — защитную, — в случае повреждения ноцицепция переключается с предотвращения повреждающей стимуляции на заживление травмированной ткани.
Как было сказано выше, ноцицептивная боль является защитной реакцией организма, способствующей выживанию индивида. При воспалении адаптивное значение боли утрачивается. Поэтому, несмотря на то, что боль при воспалении является ноцицептивной, некоторые авторы выделяют ее в самостоятельную форму.
Воспалительная боль характеризуется повышенной чувствительностью к стимулам, которые не вызывают боль, что защищает человека от дальнейшего повреждения травмированной ткани, пока восстановительный процесс не будет закончен. Исключением является боль, возникающая при хронических заболеваниях, таких как ревматоидный артрит. В таком случае нужно иметь в виду, что боль следует нормализовать, не затрагивая систему ноцицептивной боли .
Невропатическая боль, вопреки ноцицептивной, возникает при прямом повреждении или заболевании нейронов периферической или центральной нервной системы и не оказывает никакого благотворного влияния. Помимо этого, периферические невропатические болевые синдромы отличаются от центральной боли. Невропатическая боль чувствуется отлично от обычной боли и не относится ни к какому сигналу о тканевом повреждении. Она возникает спонтанно и ассоциирована с другими сенсорными дефектами. Такая боль имеет обжигающий или электрический характер и может быть комбинирована с аллодинией и/или гипералгезией. Это обычно хроническая боль и слабо поддается лечению. Причиной могут послужить несколько факторов: повреждение корешка нерва, повреждение спинного мозга, поражение головного мозга, диабетическая полинейропатия, СПИД-полинейропатия, постгерпетическая нейропатия.
Функциональная боль возникает при ненормальной реакции нервной системы. При такой боли клинически не имеют места неврологические и периферические дефекты. Её физиологической основой является гиперчувствительность сенсорной системы, что усиливает симптомы. Синдромы, соответствующие данному классу боли: фибромиалгия, синдром раздражённой толстой кишки, несердечная боль в груди, головная боль напряжения.
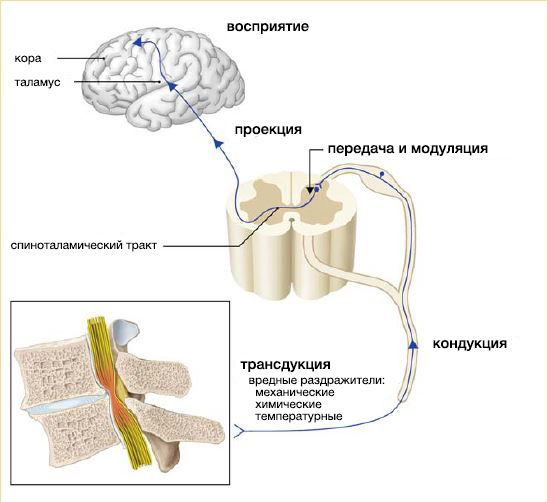
Пути проведения боли
Физиологические процессы, вовлекаемые в ощущение боли, включают:
- трансдукцию воздействующего стимула (температурного, механического или химического) в электрическую активность периферических терминалей чувствительных волокон ноцицепторов;
- кондукцию, или проведение, результирующего сенсорного воздействия на центральную терминаль ноцицепторов;
- передачу сенсорного воздействия от одного нейрона на другой и его модуляцию;
- проекцию на ствол мозга, таламус и кору;
- восприятие сенсорного воздействия в соматосенсорной коре.
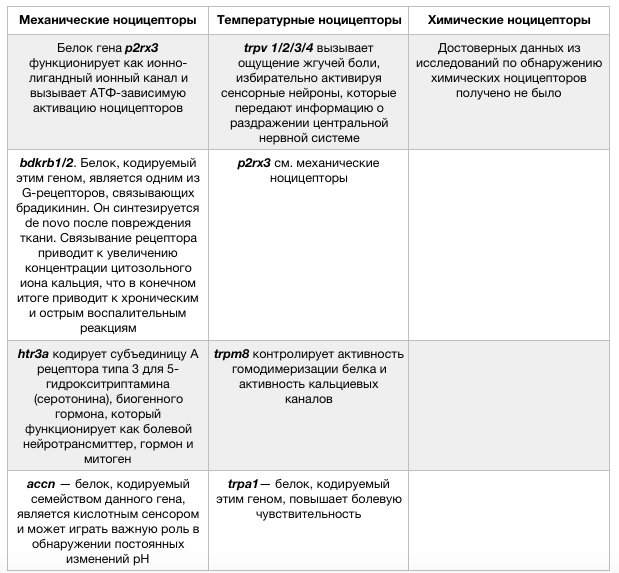
 Ноцицепция отвечает за обнаружение вредных стимулов и последующую передачу кодированной информации в головной мозг, однако боль — перцептивный процесс и полностью зависит от индивидуального восприятия данных стимулов . Ноцицепция опосредуется активацией периферических сенсорных терминалей, которые располагаются в коже, глубоких фасциях, мышцах и суставах. Такие сенсорные нейроны называются ноцицепторы. Различают три вида вредного воздействия, ориентированные на ноцицепторы: механическое давление, в трансдукции которого участвуют гены p2rx3, bdkrb 1/2, htr3a, и др., механическое повреждение (trpv4, trpc/p, accn и др.)); температурное (горячее (trpv 1/2/3/4, p2xr3 и др.) / холодное (trpm8, trpa1 и др.)); химическое.
Ноцицепция отвечает за обнаружение вредных стимулов и последующую передачу кодированной информации в головной мозг, однако боль — перцептивный процесс и полностью зависит от индивидуального восприятия данных стимулов . Ноцицепция опосредуется активацией периферических сенсорных терминалей, которые располагаются в коже, глубоких фасциях, мышцах и суставах. Такие сенсорные нейроны называются ноцицепторы. Различают три вида вредного воздействия, ориентированные на ноцицепторы: механическое давление, в трансдукции которого участвуют гены p2rx3, bdkrb 1/2, htr3a, и др., механическое повреждение (trpv4, trpc/p, accn и др.)); температурное (горячее (trpv 1/2/3/4, p2xr3 и др.) / холодное (trpm8, trpa1 и др.)); химическое.
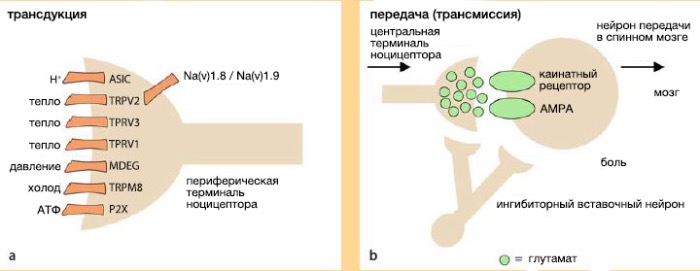
 Рассмотрим трансдукцию на примере химических раздражителей.
Рассмотрим трансдукцию на примере химических раздражителей.
Первичные сенсорные нейроны реагируют на интенсивное давление или раздражающие химикаты, но невосприимчивы к таким безвредным стимулам, как тепло и свет . Превращение вредных температурных, механических или химических раздражителей в электрическую энергию в периферических терминалях сенсорных волокон ноцицепторов называется трансдукцией.
Механическое давление, тканевая деформация или осмолярные изменения могут активировать ноцицепторы, что позволяет распознать прикосновение, сильное давление, изменения внутренних органов, костную деструкцию или отёк . Эти стимулы опосредованы механосенсорными преобразователями, такими как ионные каналы семейства mDEG (дегенерин млекопитающих) или кислоточувствительные рецепторы 2 (ASIC2).
Механическое раздражение может высвободить клеточный АТФ, активируя связанные с G-белком АТФ-рецепторы (P2Y) или АТФ-зависимые ионные каналы (P2X). Чрезмерное нагревание может быть определено ванилоидными рецепторами (TRPV1, которые ранее назывались VR1) и ванилоидным ионотропным каналом (TRPV2, или VRL1), который принадлежит к более широкому семейству каналов, регулирующих проницаемость для катионов, главным образом для Са2+ (TRP). Структура ядерной мембраны рецепторов аналогична с потенциал-зависимыми К+ или CNG-каналами . Рецептор TRPM8, отдалённый родственник TRPV1, отвечает за распознавание чрезмерного холода.
Ноцицепторы однозначно экспрессируют два потенциал-зависимых натриевых канала (Na(v)1.8 и Na(v)1.9), которые могут стать мишенью для селективных анестетиков, блокирующих только боль, оставляя ощущение безвредных стимулов.
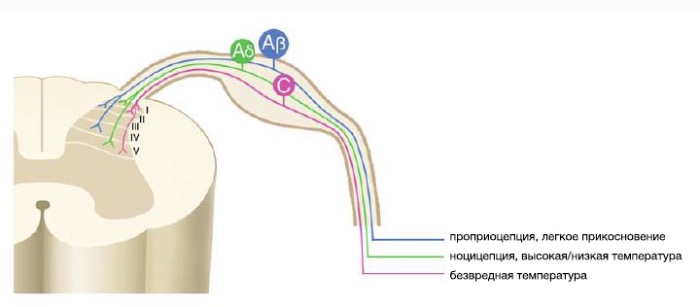
Волокна типа С демиелинизированы, их диаметр порядка 0,4-1,2 нм, скорость проведения 0,5-2,0 м/с. Они имеют термочувствительные рецепторы, механорецепторы с низким порогом чувствительности и специфические рецепторы для альгогенных веществ.
Aδ-волокна слабомиелинизированы, диаметр 2,0-6,0 нм, скорость проведения 12-30 м/с. Эти волокна подразделяются на две подгруппы. Aδ-волокна типа I представляют собой высокопороговые механорецепторы, слабо реагирующие на химические и термические раздражители. Тип II соответствует, главным образом, механотермическим рецепторам для высоких температур и интенсивного холода.
Аβ-волокна миелинизированы, диаметр составляет более 10 нм, скорость проведения 30-100 м/с. Эти волокна чувствительны к прикосновениям, легкому давлению и вибрации, а также осуществляют проприоцепцию. Их активация способствует механизмам сегментарного подавления в спинном мозге.
Активация волокон типа С и Aδ ведёт к обжигающим болевым ощущениям. При патологических состояниях признаки нейропатической боли, например, дизестезия и парестезия, могут возникнуть в результате активации волокон Aβ. Патологическое болевое ощущение может проявляться как гипералгезия, опосредуемая активацией С- и Аδ-волокнами. При патологических состояниях активация низкопороговых механорецепторов (волокон Aβ) может вызывать аллодинию (боль вследствие воздействия раздражений, обычно её не вызывающих).
Под трансмиссией (передачей) понимают синаптический переход сенсорного импульса с одного нейрона на другой. Импульс передаётся с клеток спинномозговых ганглиев на нейроны задних рогов. В конечном счёте, для продолжения прохождения афферентного сигнала в вышележащие структуры импульс должен переключиться на проекционные нейроны прямо или опосредованно через собственные.
Цитоархитектура задних рогов спиного мозга
Серое вещество спинного мозга можно разделить на десять слоев: I слой — краевая пластина — образован многими мелкими нейронами и крупными веретеновидными клетками, лежащими параллельно самой пластине; II и III слой — желатинозная субстанция; IV слой соответствует собственному ядру; V и VI (глубокие слои) составляют задние рога спинного мозга ; VII занимает значительную часть переднего рога; VIII слой расположен в вентро-медиальной части переднего рога, вокруг одной из частей IX слоя; IX не един в пространстве, его части лежат внутри VII и VIII слоев; и, наконец, X слой расположен вокруг спинального канала, и отвечает за комиссуральные (между левой и правой частями спинного мозга) и другие проприоспинальные связи.
Столбики пластинчатой формы, находящиеся в задних рогах, простираются вдоль спинного мозга . Внутри столбиков находится большое количество возбуждающих и ингибирующих вставочных нейронов второго порядка, которые получают множественные входные сигналы от окружающих структур и посылают результаты в мозг и в передние рога . Таким образом, нейронная сеть задних рогов служит в качестве ворот, контролирующих распространение ноцицептивных сигналов в более высокие области мозга.
Цитоархитектура задних рогов спинного мозга очень сложна. Упрощенно, миелинизированные низкопороговые Аβ-волокна оканчиваются в III и IV слое, в то время как слабомиелинизированные высокопороговые Aδ-волокна проходят до I и V слоя, а немиелинизированные высокопороговые С-волокна оканчиваются во II слое, однако, некоторые идут как и Aδ-волокна, до I и V слоев . В заднем роге можно выделить три различных типа нейронов, которые различаются типом афферентов их ответной реакции на ноцицептивное воздействие : специфичные ноцицептивные нейроны, мультирецепторные, или нейроны широкого динамического диапазона (WDR нейроны), и неноцицептивные нейроны. Остановимся подробнее на каждом типе нейронов.
Специфичные ноцицептивные нейроны располагаются в желатинозной субстанции, но также могут физиологически локализоваться в V и VI слоях. Они активируются исключительно высокоинтенсивным вредным стимулом от С и Aδ-волокон.
Мультирецепторные нейроны, или нейроны широкого динамического диапазона (WDR нейроны), отвечают на температурное, механическое и химическое раздражение, переданное через С-, Aδ- и Аβ-волокна. Эти нейроны в меньшей степени располагаются в вентральных рогах спинного мозга. WDR нейроны представляют собой слияние от кожного, мышечного и висцерального входов. Их первостепенной задачей является кодирование интенсивности раздражения . Кроме того, эти нейроны участвуют в опосредованных C-волокнами процессах сенсибилизации и усиления длительной боли.
Неноцицептивные нейроны (N-NOC) активируются безвредными раздражителями, такими как механическое, температурное или проприоцептивное воздействие низкой интенсивности, переданное Aδ- и Аβ-волокнами, и встречаются во II, III и IV слоях . Эти нейроны косвенно действуют в сегментарных супрессионных механизмах . Разные типы нейронов соединены возбуждающими и тормозными вставочными нейронами второго порядка, которые принимают сигнал от других колонн и передают информацию и импульсы в головной мозг . После модуляции и модификации ноцицептивных стимулов в заднем роге информация передаётся в ЦНС.
Афференты нейронов задних рогов спинного мозга образуют так называемые позвоночные пути, которые передают ноцицептивную информацию в ЦНС. Пластичность, или модифицируемость, синаптического переноса в рогах спинного мозга является ключевой особенностью его функции и является неотъемлемой частью генерации боли и её гиперчувствительности . Основные синапсы, ответственные за передачу, расположены в дорсальном роге спинного мозга в I слое (краевая зона) и во II слое (желатинозная субстанция). Импульсы передаются в таламус — основную область интеграции информации. Трансфер ноцицептивных стимулов может ингибироваться нисходящими путями ствола мозга, среднего мозга и коллатеральными влияниями в спинном роге. Теперь перейдем к одному из самых важных и сложных этапов проведения боли, который включает в себя взаимодействие со всем организмом человека — модуляции.
Модуляция чувствительных сигналов
По ходу передачи периферических ноцицептивных сигналов в головной мозг они подвергаются различным влияниям нисходящих путей . Изучено множество нейротрансмиттеров, контролирующих такое воздействие.
Модуляция — это процесс, в котором передаваемая информация подвергается изменениям (модуляции) прежде, чем окажется в ЦНС. Существуют два типа механизмов модуляции ноцицептивных импульсов: тормозные и возбуждающие.
Тормозные механизмы могут происходить от локальных (сегментных) ингибирующих вставочных нейронов или от нисходящих антиноцицептивных путей. Большинство локальных ингибиторных нейронов в спинном мозге высвобождают глицин и/или γ-аминомасляную кислоту (ГАМК). Нисходящие тормозные пути берут начало на уровне коры и таламуса и спускаются через ствол головного мозга (околоводопроводное серое вещество) и задний столб, заканчиваясь в заднем роге спинного мозга. Эти нисходящие пути модулируют ноцицептивную передачу путем высвобождения серотонина и/или норадреналина, а тормозное воздействие может быть постсимпатическим и пресимпатическим. Постсимпатическое воздействие приводит к гиперполяризации клеточной мембраны и/или активации шунта изоляции, которая препятствует распространению возбуждающих постсинаптических потенциалов вдоль дендрита нейронов. Пресинаптическое воздействие происходит в аксо-аксональных синапсах ГАМКергических нейронов с первичными сенсорными нервными терминалями.
 Возбуждающие механизмы. Глутамат — возбуждающий нейротрансмиттер, высвобождающийся первичными афферентными волокнами, играет ключевую роль в механизме спинальной передачи боли . Синаптически высвобожденный глутамат действует на каинатные и AMPA (α-амино-3-гидрокси-5-метил-4-изоксазол-пропионовая кислота) рецепторы, отвечая за быструю синаптическую передачу в первом синапсе заднего рога. Безвредные стимулы приводят к стабильным AMPA-опосредованным синаптическим сигналам, которые в конечном итоге воспринимаются как преходящая локализованная боль. Глутамат также воздействует на N-метил-D-аспартат (NMDA) рецепторы, однако в условиях покоя этот рецептор заблокирован внеклеточными ионами магния. Деполяризация постсинаптического нейрона, например, через интенсивную активацию AMPA-рецептора, снимает этот блок. Помимо этого, уменьшить чувствительность NMDA-рецептора к ионам магния могут активаторы протеинкиназы С, что, возможно, будет способствовать гиперчувствительности спинного мозга и усилению периферических импульсов. Активация NMDA-рецептора также приведёт к входу кальция, что является ключевым событием в генерации усиления синаптической передачи между двумя нейронами, сохраняющейся на протяжении длительного времени после воздействия на синаптический проводящий путь (долговременная потенциация или LTP). Вдобавок ко всему выше сказанному, Са активирует различные ферменты, такие как NO-синтазу, фосфолипазы , которые тоже влияют на проведение боли.
Возбуждающие механизмы. Глутамат — возбуждающий нейротрансмиттер, высвобождающийся первичными афферентными волокнами, играет ключевую роль в механизме спинальной передачи боли . Синаптически высвобожденный глутамат действует на каинатные и AMPA (α-амино-3-гидрокси-5-метил-4-изоксазол-пропионовая кислота) рецепторы, отвечая за быструю синаптическую передачу в первом синапсе заднего рога. Безвредные стимулы приводят к стабильным AMPA-опосредованным синаптическим сигналам, которые в конечном итоге воспринимаются как преходящая локализованная боль. Глутамат также воздействует на N-метил-D-аспартат (NMDA) рецепторы, однако в условиях покоя этот рецептор заблокирован внеклеточными ионами магния. Деполяризация постсинаптического нейрона, например, через интенсивную активацию AMPA-рецептора, снимает этот блок. Помимо этого, уменьшить чувствительность NMDA-рецептора к ионам магния могут активаторы протеинкиназы С, что, возможно, будет способствовать гиперчувствительности спинного мозга и усилению периферических импульсов. Активация NMDA-рецептора также приведёт к входу кальция, что является ключевым событием в генерации усиления синаптической передачи между двумя нейронами, сохраняющейся на протяжении длительного времени после воздействия на синаптический проводящий путь (долговременная потенциация или LTP). Вдобавок ко всему выше сказанному, Са активирует различные ферменты, такие как NO-синтазу, фосфолипазы , которые тоже влияют на проведение боли.
Часто повторяющаяся стимуляция волокон C приводит к увеличению ответной реакции, даже несмотря на то, что амплитуда входного сигнала остаётся неизменной. Это независящее от деятельности явление, известное как «зависание», несёт ответственность за усиление боли, возникающей в результате повторяющегося раздражения кожи слишком высокой температурой. Далее наступает этап, который можно представить в виде передвижения боли по своеобразным рельсам в верхние отделы — проекция боли.
Проекция боли
После передачи и модуляции боли внутри дорсального рога, ноцицептивная информация проецируется в супраспинальные структуры через афферентные чувствительные волокна. Они проникают в спинной мозг через зону вхождения заднего корешка (зона Редлиха-Оберштейнера), а затем отдают многочисленные коллатерали, образующие в спинном мозге синаптические контакты с другими нейронами. Все пучки волокон, каждому из которых в спинном мозге отведено свое место, относятся к определенному виду чувствительности. Эти пучки можно разделить на несколько трактов со специальными функциями:
- Спиноталамический тракт участвует в узнавании боли, её аффективных и мотивационно-аффективных аспектов;
- Спинно-ретикулярный тракт участвует в мотивационно-аффективных аспектах и нейровегетативных реакциях на боль;
- Спиномезенцефалический тракт участвует в соматосенсорной обработке, активации нисходящей аналгезии, вызывает аверсивное поведение в ответ на ноцицептивные стимулы, а также вызывает вегетативные, сердечно-сосудистые, мотивационные и аффективные реакции;
- Спинопарабрахиальный тракт участвует в вегетативной, мотивационной, аффективной регуляции и в нейроэндокринных реакциях на боль;
- Спиногипоталамический тракт участвует в нейроэндокринных автономных, мотивационных, аффективных и тревожных ответах на соматическую и висцеральной боль;
- Спиноцервикальный тракт участвует в узнавании и мотивационно-аффективных и вегетативных реакциях на боль, а так же играет роль в сенсорной интеграции и модуляции афферентных сигналов;
- Постсинаптические пути позвоночного столба, участвующие в сенсорно-дискриминационных компонентах и мотивационно-аффективных аспектах боли.
Стоит заметить, что по мере прохождения афферентных волокон через зону вхождения корешка и проникновения в задний рог, их миелиновая оболочка заметно истончается. Более того, от периферических отделов нервной системы к центральным меняется и тип миелина, поскольку там миелин образуется уже не леммоцитами, а олигодендроцитами.
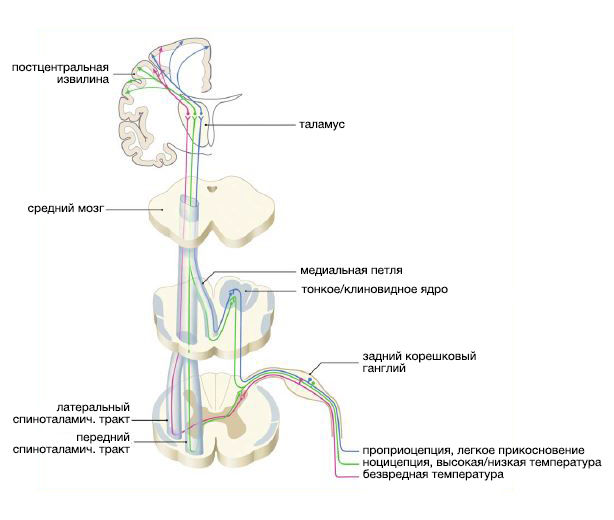 Афферентные пути
Афферентные пути
В данном обзоре мы уделим внимание восприятию боли и концепции её лечения, а подробному описанию хода и состава волокон трактов будет посвящена отдельная статья.
Восприятие боли
Здесь сразу стоит уточнить, что боль в каждом случае воспринимается субъективно и зависит от великого множества причин, таких как генетический фон, порог болевого ощущения, индивидуальный интервал выносливости, личностные качества и т.п., но процесс восприятия всегда одинаков. Пути спинальной проекции идут в направлении ретикулярной формации ствола мозга и окружающих её ядер до схождения в таламусе, главной структуре для приема, и интеграции ноцицептивных стимулов перед передачей их в соматосенсорную кору. Однако лишь незначительная часть всех сенсорных сигналов из спинного мозга поступает в таламус из-за отбора их на предыдущих этапах (локальная обработка, модуляция и т.д.) . Соматосенсорная кора, в свою очередь, проецирует их в смежные области коры, преимущественно лимбической системы. В лимбическую систему входят :
- поясная извилина (поведение и эмоции);
- амигдала (обусловленный страх и беспокойство);
- гиппокамп (память);
- гипоталамус (симпатическая вегетативная активность);
- голубое пятно (пробуждение, бдительность, поведение);
- части околоводопроводного серого вещества (реакция «бей или беги», стресс-индуцированная анальгезия).
Проекции от околоводопроводного серого вещества играют роль в контроле антиноцицептивных и вегетативных реакций на ноцицептивные стимулы .
Нейропластичность
Постоянная боль — это не просто пролонгирование острой (ноцицептивной) боли, а результат процесса альтерации в составляющих частях путей её проведения. Повреждение периферической ткани или повреждение нерва могут приводить к патологическому состоянию, в котором происходит снижение болевого порога (аллодиния), усиление реакции на вредные стимулы (гипералгезия), увеличение продолжительности реакции на кратковременную стимуляцию (постоянная боль) и распространение боли и гипералгезии на неповрежденную ткань (боль в спине и вторичная гипералгезия) . Эти изменения в путях проведения боли называют нейропластичностью.
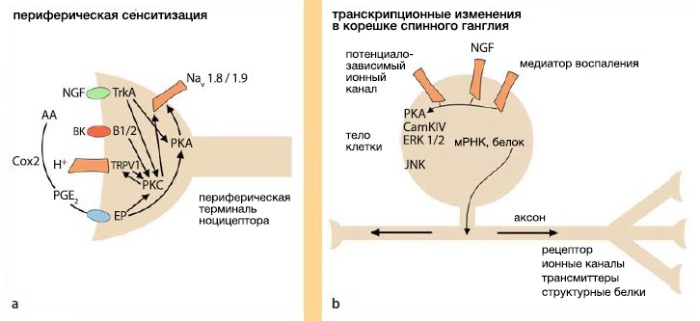
Периферическая сенсибилизация
Повреждение ткани приводит к высвобождению медиаторов воспаления, включая ионы (H+, K+), брадикинин, гистамин, 5-гидрокситриптамин, (5-HT), АТФ и оксид азота (NO). Травма ткани активирует путь арахидоновой кислоты, что приводит к образованию простаноидов и лейкотриенов . Воспалительные медиаторы также выделяются из привлекаемых клеток, таких как тучные клетки, фибробласты, нейтрофилы и тромбоциты. Повреждение тканей и воспаление приводят к понижению рН, что усиливает болевые ощущения, активируя и сенсибилизируя ваниллоидный рецептор 1 (TRPV1) . Воспалительные медиаторы, например, простагландин E2, брадикинин и фактор роста нервов (NGF), активируют в периферических терминалях внутриклеточные протеинкиназы А и С, которые фосфорилируют TRPV1 и устойчивые к тетрадотоксину (TTXr) рецепторы натриевых каналов (Na(v)1.8, Na(v)1.9) для повышения возбудимости . Эти механизмы способствуют сенсибилизации периферических терминалей, что приводит к болевой гиперчувствительности.
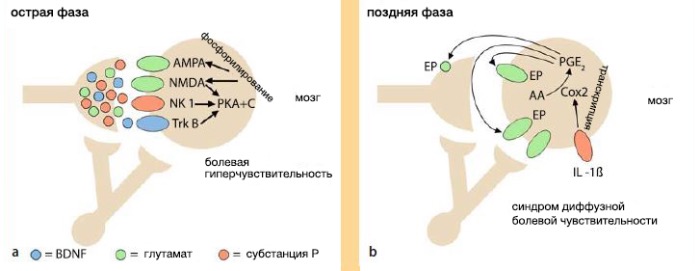
Центральная сенсибилизация
Центральная сенсибилизация представляет собой форму синаптической пластичности, которая усиливает и облегчает синаптическую передачу от центрального ноцицептора к нейронам спинного рога . Во время процесса ноцицепции высвобождение глутамата преимущественно действует на каинатные и АМРА-рецепторы внутри дорсального рога. Интенсивная стимуляция ноцицепторов (например, травма позвоночника) высвобождает трансмиттеры (нейротрофический фактор мозга (BDNF), субстанция P, глутамат), которые действуют на многочисленные рецепторы спинного рога, т.е. AMPA, NMDA, NK1 и TrkB . В ранней фазе центральной сенсибилизации также активируются внутриклеточные киназы, которые фосфорилируют рецепторы ионных каналов. Этот эффект повышает чувствительность к глутамату путем удаления блока для Mg2+ канала NMDA, приводящего к гиперчувствительности спинного мозга и усилению периферических входов.
Дезингибирование (растормаживание)
Афферентные ноцицептивные сигналы от периферии к мозгу модулируются хорошо сбалансированным взаимодействием возбуждающих и тормозных нейронов . Потеря торможения, то есть дезингибирование нейронов дорсальных рогов, является ключевым элементом стойкой воспалительной нейропатической боли . Ингибиторные механизмы в спинном мозге опосредованы нейротрансмиттерами глицином и ГАМК. Экспрессия PGE2 во время воспаления приводит к A-зависимому фосфорилированию протеинкиназы, которое ингибирует рецепторы глицина. К этому этапу нейроны спинного рога освобождаются от глицинергической нейротрансмиссии. Кроме того, известно, что частичное повреждение нерва уменьшает уровень ГАМК-синтезирующего фермента декарбоксилазы глутаминовой кислоты (GAD) и индуцирует нейрональный апоптоз. Оба эти механизма могут снижать пресинаптические уровни ГАМК и способствовать функциональной недостаточности ГАМК-ергической передачи в поверхностных слоях заднего рога. Однако для развития тепловой гипералгезии в модели хронического констриктивного повреждения (CCI), как причине нейропатической боли, значительная потеря ГАМК-ергических или глицинергических нейронов не обязательна.
Дополнительные механизмы, вовлеченные в нейропластичность и приводящие к обработке патологической боли, включают глиальные изменения в спинном мозге и медуллярное нисходящее растормаживание. Подобно иммунным клеткам, реагирующим на вирусы и бактерии, глия спинного мозга (микроглия и астроциты) может усиливать боль, высвобождая провоспалительные цитокины . Этот процесс также активируется некоторыми сенсорными сигналами, поступающими с периферии, например, повреждением нервного корешка. Травма нервного корешка и/или воспаление могут приводить к постоянному поступлению болевых сигналов и длительной активации нисходящих модуляторных путей, которые облегчают передачу боли.
Основные концепции лечения боли
В этой части работы будет рассмотрено лечение боли на примере боли в нижнем отделе позвоночника, как одного из наиболее часто встречающихся вариантов болевого синдрома.
Фармакологическое лечение
Системное фармакологическое лечение острой и хронической боли является достаточно сложным и очень важным процессом. Его по праву называют краеугольным камнем в науке о боли . Трехступенчатая терапия обезболивания, разработанная ВОЗ в 1986 году, первоначально предназначалась для лечения боли при онкологическом заболевании. Вскоре она также показала эффективность и при других болевых расстройствах, например, боли в спине. Первым шагом такая терапия подразумевает применение слабого анальгетика с поэтапным увеличением дозы до полного исчезновения боли. В случаях же сильной боли может потребоваться немедленное применение наркотических опиоидных анальгетиков (стратифицированная терапия, шаг 3). Появляется все больше свидетельств того, что острые болезненные переживания могут влиять на продолжительность боли даже после заживления тканей. Растущее понимание нейробиологии боли побудило создать агрессивный, мультимодальный, превентивный подход к лечению острой боли, чтобы предотвратить её переход в хроническую форму.
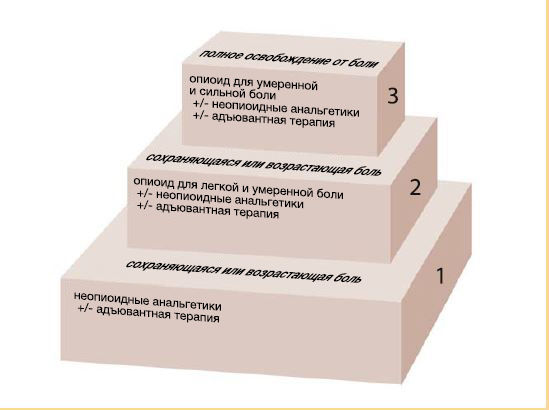 Ненаркотические анальгетики
Ненаркотические анальгетики
Парацетамол является наиболее часто используемыми ненаркотическими анальгетиками. Несмотря на то, что представитель группы ненаркотических анальгетиков — парацетамол (ацетаминофен) — известен уже более сотни лет, механизмы его антиноцицептивного эффекта остаются спорными. По известным данным, парацетамол вызывает слабое периферическое и центральное ингибирование циклооксигеназы (ЦОГ) . Считается, что анальгезирующее действие парацетамола связано с увеличением болевого порога посредством центрального ингибирования простагландинов. Антагонисты NMDA-рецепторов — это мощные анальгетики, которые препятствуют трансмиссии в первичных афферентных путях боли у NMDA-рецепторов. Прототипом антагонистов NMDA-рецепторов является кетамин, который эффективен при невропатических и других хронических болевых состояниях.
Нестероидные противовоспалительные препараты
НПВС являются фундаментом лечения боли воспалительной природы. Основным механизмом действия нестероидных противовоспалительных препаратов (НПВС) является ингибирование синтеза простагландинов путем блокирования циклооксигеназы (ЦОГ), которая катализирует биотрансформацию арахидоновой кислоты в простагландины. В большинстве тканей ЦОГ-1 экспрессируется конститутивно, тогда как экспрессия ЦОГ-2 индуцируется во многих типах клеток в результате воспаления . Продукты ЦОГ-1 и ЦОГ-2, в частности, простагландин Е2 и I2, вызывают воспалительные изменения и действуют непосредственно на сенсорные нервные окончания . Неселективные ингибиторы ЦОГ (например, аспирин, ибупрофен, напроксен, диклофенак, пироксикам) ингибируют обе изоформы ЦОГ. Однако ингибирование ЦОГ-1 имеет один недостаток: он также предотвращает синтез ПГ, чья функция — защита тканей. После открытия изоферментов ЦОГ были разработаны селективные ингибиторы ЦОГ-2. Однако недавно они были тщательно исследованы (например, целекоксиб, рофекоксиб, вальдекоксиб) по подозрению о потенциально серьезных побочных эффектах.
Наркотические анальгетики
Опиоиды являются основой терапии острой боли. Они включают все эндогенные и экзогенные соединения, которые обладают морфиноподобными анальгетическими свойствами. Среди наиболее часто используемых опиоидов — морфин, гидроморфон, трамадол, метадон, оксикодон, оксиморфон и фентанил. Эти препараты остаются основой для лечения тяжелых острых болей. Существуют споры об их эффективности и безопасности при длительном использовании. Впрочем, недавний систематический обзор показал, что кратковременное применение опиоидов имеет положительный эффект как при нейропатической, так и при боли в опорно-двигательного аппарата. Тем не менее, выводы о толерантности и зависимости были невозможны из-за небольшого числа пациентов, долгосрочно принимающих препарат, что не позволяет делать выводы о лечении хронической боли.
Адъювантная терапия
ВОЗ рекомендовала добавлять адъювантные лекарственные средства для облегчения связанных с болью страхов и тревоги , а также чтобы усилить основной обезболивающий эффект. Можно дифференцировать несколько категорий адъювантных препаратов:
- Антидепрессанты;
- Противосудорожные средства;
- Анксиолитики;
- Миорелаксанты;
- Лекарства для сна.
Трициклические антидепрессанты (например, амитриптилин, дезипрамин, нортриптилин) имеют долгую историю использования при нейропатической боли и действуют главным образом путем усиления адренергической стимуляции α2-адренорецепторов. Некоторые также обладают блокирующей NMDA-рецептор активностью. Обоснование их использования при хронической боли в пояснице основано на частом сосуществовании боли и депрессии, седативном эффекте препаратов (улучшении сна) и предполагаемом анальгетическом эффекте в более низких дозах. Тем не менее, существуют противоречивые данные о том, что антидепрессанты эффективны при болях в пояснице в краткосрочной и среднесрочной перспективе.
Противосудорожные препараты чрезвычайно полезны при нейропатической боли . Их эффективность при лечении невропатических и центральных болевых состояний заключается в неселективном блокирующем Na+ - каналы действии. До недавнего времени первое поколение противосудорожных средств (например, фенитоин, карбамазепин и вальпроевая кислота) применяли для лечения нейропатической боли. Однако новые противоэпилептические средства, включая габапентин и прегабалин, быстро становятся препаратами первой линии.
Селективные ингибиторы обратного захвата серотонина (например, флуоксетин, пароксетин) часто используются для лечения тревожных расстройств. Однако терапевтические эффекты наблюдаются не сразу из-за медленного начала действия (2-4 недели). Бензодиазепины используются для лечения острых тревожных состояний и служат в качестве премедикации перед хирургическим вмешательством для снижения стресса и вероятности мышечного спазма.
Мышечные релаксанты оказывают непрямое периферическое воздействие на мышечный спазм, а центральное воздействие на нервную систему. Бензодиазепины (например, диазепам) являются седативными средствами и проявляют аддиктивный потенциал, а также могут вызвать синдром отмены. Баклофен централизует передачу сигналов, опосредованных GABAb-рецептором, тогда как тизанидин является центрально-действующим α2-адренергическим агонистом и уменьшает выделение возбуждающих нейротрансмиттеров и ингибирует спинальные рефлексы . Есть убедительные доказательства того, что оральные небензодиазепиновые препараты являются более эффективными для пациентов с острой болью в спине чем плацебо. Разница проявляется в кратковременном облегчении боли, эффективности и улучшении физических возможностей пациентов. Тем не менее, нет достаточного количества доказательств, чтобы быть уверенным в аналогичных эффектах при хронической боли в спине.
Лекарства, стимулирующие сон, полезны в качестве адъювантной терапии из-за высокой корреляции бессонницы, депрессии и боли . Поэтому соответствующее лечение боли также помогает при бессоннице. Традиционно, антидепрессанты используются в силу их седативного эффекта. Бензодиазепины следует использовать только для краткосрочного лечения бессонницы из-за известных побочных эффектов, таких как чрезмерная седация («утреннее похмелье»), зависимость, абстинентный синдром. Новые препараты — агонисты рецептора омега-1 (например, золпидем, залеплон) — сводят к минимуму утреннее похмелье и симптомы отмены, а также имеют более короткий период полураспада .
Нефармакологическое лечение
Хорошо известно, что постельный режим более 5 дней не рекомендуется при острой боли в спине. Имеются противоречивые данные об эффективности разнообразных упражнений для пациентов с хронической болью. В то же время существуют данные о влиянии лечебной физкультуры на острую боль. Упражнения по меньшей мере столь же эффективны, сколько и другие консервативные вмешательства при хронической боли. Спинальные манипуляции не являются более эффективными в краткосрочной и долгосрочной перспективе по сравнению с другими традиционно поддерживаемыми вариантами терапии, такими как общая терапия, физическая нагрузка и лечебная физкультура.
Биопсихологическое вмешательство
Начиная с введения Мелзаком и Уоллом “теории ворот”, или теории блокаторов боли, наше понимание того, как психосоциальные факторы могут модулировать сигнал боли, значительно увеличилось. Кроме того, оно было сформировано и другой вехой. В конце 1970-х годов Энгель осознал, что доминирующая биомедицинская модель не оставляет места в ее рамках для социальных, психологических и поведенческих аспектов болезни. Так, он предложил биопсихосоциальную модель, которая включала не только физиологические, но также психологические и социальные факторы, позволяющие более полно понять боль. Эти два теоретических аспекта привели к разработке принципиально новых подходов к лечению, т.е. поведенческому и когнитивно-поведенческому методам, которые выходят за рамки биомедицинских измерений. Обоснованием такого подхода является изменение диапазона физических, психологических и социальных компонентов боли.
При постоянных болевых расстройствах фактическое повреждение тканей со временем почти всегда исчезает, а режим, содействующий заживлению, более не требуется. Совет оставаться максимально активными — это очень важный совет, который следует давать пациентам. Есть доказательства того, что, по крайней мере, в краткосрочной перспективе, занятость позволяет уменьшить интенсивность боли. Fordyce и его коллеги также указали, что боль ощущается не так сильно, если вы чем-то заняты.
Несмотря на доказательства того, что когнитивно-респондентное и интенсивное мультидисциплинарное лечение является эффективными для краткосрочного уменьшения боли при хроническом болевом синдроме, до сих пор нет доказательств того, что какое-либо из подобных вмешательств оказывают долгосрочное воздействие на боль в пояснице.
Хирургическое лечение
Хирургическое лечение при постоянной неспецифической боли не имеет достаточной доказательной базы . До сих пор отсутствуют убедительные доказательства превосходства таких методов над когнитивно-поведенческим лечением и физическими упражнениями. Аналогичным образом, нет других инвазивных вмешательств (например, спинальная инъекция, стимуляция спинного мозга, интратекальные насосы) для лечения хронической боли в пояснице, отличной от грыжи диска, спинного стеноза и спондилолистеза (при которых проводится хирургическая декомпрессия и стабилизация).
Заключение
Так, боль, проходя через все физиологические этапы: трансдукцию, кондукцию, передачу, модуляцию, проекцию и восприятие сенсорного воздействия, следуя по различным волокнам, взаимодействуя со множеством нейротрансмиттеров, испытывая на себе всю мощь нейропластичности, усиливаясь и ослабляясь, незамедлительно дает о себе знать. Как известно, в ответ на боль (а также при приеме пищи) в организме выделяются большое количество разных веществ, одними из которых являются эндогенные опиоидные пептиды, а именно энкефалины и эндорфины. Физиологически, эндорфины и энкефалины обладают сильнейшим обезболивающим, противошоковым и антистрессовым действием, они понижают аппетит и уменьшают чувствительность отдельных отделов центральной нервной системы. Человек становится "слеп от счастья" — если говорить утрировано. В момент получения травмы организм высвобождает большое количество опиоидов, которые действовали до поры до времени, однако вскоре их действие заканчивается и наступает момент, когда человек начинает чувствовать боль.
Остается загадкой, знал ли майор Пэйн о физиологии эндогенных опиоидных веществ, но, сломав палец солдату, он заставил выделить организм новую порцию энкефалинов и эндорфинов, что на какое-то время должно было помочь солдату не чувствовать боль. Как говорится,“ноу Пэйн — ноу гейн”. Конечно, сегодня методы лечения острой и хронической боли шагнули далеко вперед и их совершенствование продолжается, в основе которого лежат новые достижения фундаментальных наук.
Французы — большие мастера крылатых фраз. “Скажи мне кто твой друг и я скажу кто ты”, “всему свое время”, “аппетит приходит во время еды” — все это французский гений мысли. И именно французским выражением я бы хотел закончить этот обзор. Дорогие читатели, не забывайте — “мы живы до тех пор, пока нам больно”.
Источники
-
Ahmadi S. et al. PGE2 selectively blocks inhibitory glycinergic neurotransmission onto rat superficial dorsal horn neurons //Nature neuroscience. – 2002. – Т. 5. – №. 1. – С. 34-40.
-
Almeida T. F., Roizenblatt S., Tufik S. Afferent pain pathways: a neuroanatomical review //Brain research. – 2004. – Т. 1000. – №. 1. – С. 40-56.
-
Gaston-Johansson F. et al. Similarities in pain descriptions of four different ethnic-culture groups //Journal of pain and symptom management. – 1990. – Т. 5. – №. 2. – С. 94-100.
-
Foulkes T., Wood J. N. Pain genes //PLoS genetics. – 2008. – Т. 4. – №. 7. – С. e1000086.
-
Besson J. M. The neurobiology of pain //The Lancet. – 1999. – Т. 353. – №. 9164. – С. 1610-1615.
-
Kehlet H., Dahl J. B. The value of" multimodal" or" balanced analgesia" in postoperative pain treatment //Anesthesia & Analgesia. – 1993. – Т. 77. – №. 5. – С. 1048-1056.
-
Costigan M., Woolf C. J. Pain: molecular mechanisms //The Journal of Pain. – 2000. – Т. 1. – №. 3. – С. 35-44.
-
Crofford L. J. et al. Cardiovascular effects of selective COX-2 inhibition: is there a class effect? The International COX-2 Study Group //The Journal of rheumatology. – 2006. – Т. 33. – №. 7. – С. 1403-1408.
-
Deyo R. A., Nachemson A., Mirza S. K. Spinal-fusion surgery—the case for restraint //The Spine Journal. – 2004. – Т. 4. – №. 5. – С. S138-S142.
-
Dubner R., Hargreaves K. M. The neurobiology of pain and its modulation //The Clinical journal of pain. – 1988. – Т. 5. – С. S1-4; discussion S4-6.
-
Dworkin R. H. et al. Advances in neuropathic pain: diagnosis, mechanisms, and treatment recommendations //Archives of neurology. – 2003. – Т. 60. – №. 11. – С. 1524-1534.
-
Ekman E. F., Koman L. A. Acute pain following musculoskeletal injuries and orthopaedic surgery: mechanisms and management //JBJS. – 2004. – Т. 86. – №. 6. – С. 1316-1327.
-
Elliott A. M. et al. The course of chronic pain in the community: results of a 4-year follow-up study //Pain. – 2002. – Т. 99. – №. 1. – С. 299-307.
-
Engel G. L. The need for a new medical model: a challenge for biomedicine //Holistic Medicine. – 1989. – Т. 4. – №. 1. – С. 37-53.
-
Fey S. G., Fordyce W. E. Behavioral rehabilitation of the chronic pain patient //Annual review of rehabilitation. – 1982. – Т. 3. – С. 32-63.
-
Fordyce W. E. Behavioral factors in pain //Neurosurgery Clinics of North America. – 1991. – Т. 2. – №. 4. – С. 749-759.
-
Freeman R. The treatment of neuropathic pain //CNS spectrums. – 2005. – Т. 10. – №. 9. – С. 698-706.
-
Fürst S. Transmitters involved in antinociception in the spinal cord //Brain research bulletin. – 1999. – Т. 48. – №. 2. – С. 129-141.
-
Gillespie P. G., Walker R. G. Molecular basis of mechanosensory transduction //Nature. – 2001. – Т. 413. – №. 6852. – С. 194.
-
Gottschalk A., Wu C. L., Ochroch E. A. Current treatment options for acute pain //Expert opinion on pharmacotherapy. – 2002. – Т. 3. – №. 11. – С. 1599-1611.
-
Heim H. et al. Selective COX-2 inhibitors and risk of thromboembolic events-regulatory aspects //THROMBOSIS AND HAEMOSTASIS-STUTTGART-. – 2006. – Т. 96. – №. 4. – С. 423.
-
Hellwig N. et al. TRPV1 acts as proton channel to induce acidification in nociceptive neurons //Journal of Biological Chemistry. – 2004. – Т. 279. – №. 33. – С. 34553-34561.
-
Merskey H. E. Classification of chronic pain: descriptions of chronic pain syndromes and definitions of pain terms //Pain. – 1986.
-
Ji R. R. et al. Nociceptive-specific activation of ERK in spinal neurons contributes to pain hypersensitivity //Nature neuroscience. – 1999. – Т. 2. – №. 12.
-
Ji R. R. et al. ERK MAP kinase activation in superficial spinal cord neurons induces prodynorphin and NK-1 upregulation and contributes to persistent inflammatory pain hypersensitivity //Journal of Neuroscience. – 2002. – Т. 22. – №. 2. – С. 478-485.
-
Jin S. X. et al. p38 mitogen-activated protein kinase is activated after a spinal nerve ligation in spinal cord microglia and dorsal root ganglion neurons and contributes to the generation of neuropathic pain //Journal of Neuroscience. – 2003. – Т. 23. – №. 10. – С. 4017-4022.
-
Julius D., Basbaum A. I. Molecular mechanisms of nociception //Nature. – 2001. – Т. 413. – №. 6852. – С. 203.
-
Kalso E. et al. Opioids in chronic non-cancer pain: systematic review of efficacy and safety //Pain. – 2004. – Т. 112. – №. 3. – С. 372-380.
-
Karani R., Meier D. E. Systemic pharmacologic postoperative pain management in the geriatric orthopaedic patient //Clinical orthopaedics and related research. – 2004. – Т. 425. – С. 26-34.
-
Kehlet H., Jensen T. S., Woolf C. J. Persistent postsurgical pain: risk factors and prevention //The Lancet. – 2006. – Т. 367. – №. 9522. – С. 1618-1625.
-
Kidd B. L. What are the mechanisms of regional musculoskeletal pain? //Best Practice & Research Clinical Rheumatology. – 1999. – Т. 13. – №. 2. – С. 217-230.
-
Kidd B. L., Urban L. A. Mechanisms of inflammatory pain //British journal of anaesthesia. – 2001. – Т. 87. – №. 1. – С. 3-11.
-
Kiefer W., Dannhardt G. COX-2 inhibition and the control of pain //Current opinion in investigational drugs (London, England: 2000). – 2002. – Т. 3. – №. 9. – С. 1348-1358.
-
Lin S. Y. et al. BDNF acutely increases tyrosine phosphorylation of the NMDA receptor subunit 2B in cortical and hippocampal postsynaptic densities //Molecular brain research. – 1998. – Т. 55. – №. 1. – С. 20-27.
-
Lindström I. et al. The effect of graded activity on patients with subacute low back pain: a randomized prospective clinical study with an operant-conditioning behavioral approach //Physical therapy. – 1992. – Т. 72. – №. 4. – С. 279-290.
-
MacPherson R. D. The pharmacological basis of contemporary pain management //Pharmacology & therapeutics. – 2000. – Т. 88. – №. 2. – С. 163-185.
-
MacPherson R. D. New directions in pain management //Drugs Today. – 2002. – Т. 38. – №. 2. – С. 135-145.
-
Mannion R. J. et al. Neurotrophins: peripherally and centrally acting modulators of tactile stimulus-induced inflammatory pain hypersensitivity //Proceedings of the National Academy of Sciences. – 1999. – Т. 96. – №. 16. – С. 9385-9390.
-
Maxwell S. R. J. et al. Selectivity of NSAIDs for COX‐2 and cardiovascular outcome //British journal of clinical pharmacology. – 2006. – Т. 62. – №. 2. – С. 243-245.
-
McKemy D. D. How cold is it? TRPM8 and TRPA1 in the molecular logic of cold sensation //Molecular pain. – 2005. – Т. 1. – №. 1. – С. 16.
-
Melzack R., Wall P. D. Pain mechanisms: a new theory //Survey of Anesthesiology. – 1967. – Т. 11. – №. 2. – С. 89-90.
-
Millan M. J. The induction of pain: an integrative review //Progress in neurobiology. – 1999. – Т. 57. – №. 1. – С. 1-164.
-
Moore K. A. et al. Partial peripheral nerve injury promotes a selective loss of GABAergic inhibition in the superficial dorsal horn of the spinal cord //Journal of Neuroscience. – 2002. – Т. 22. – №. 15. – С. 6724-6731.
-
Moulin D. E. Systemic drug treatment for chronic musculoskeletal pain //The Clinical journal of pain. – 2001. – Т. 17. – №. 4. – С. S86-S93.
-
Muir III W. W., Woolf C. J. Mechanisms of pain and their therapeutic implications //Journal of the American Veterinary Medical Association. – 2001. – Т. 219. – №. 10. – С. 1346-1356.
-
КОЛОКОЛОВ О. В., СИТКАЛИ И. В., КОЛОКОЛОВА А. М. Ноцицептивная боль в практике невролога: алгоритмы диагностики, адекватность и безопасность терапии //Русский медицинский журнал. – 2015. – Т. 23. – №. 12.
-
Nakamura F., Strittmatter S. M. P2Y1 purinergic receptors in sensory neurons: contribution to touch-induced impulse generation //Proceedings of the National Academy of Sciences. – 1996. – Т. 93. – №. 19. – С. 10465-10470.
-
Obata K., Noguchi K. MAPK activation in nociceptive neurons and pain hypersensitivity //Life sciences. – 2004. – Т. 74. – №. 21. – С. 2643-2653.
-
Peier A. M. et al. A TRP channel that senses cold stimuli and menthol //Cell. – 2002. – Т. 108. – №. 5. – С. 705-715.
-
Polatin P. B., Dersh J. Psychotropic medication in chronic spinal disorders //The Spine Journal. – 2004. – Т. 4. – №. 4. – С. 436-450.
-
Polgár E. et al. Selective loss of spinal GABAergic or glycinergic neurons is not necessary for development of thermal hyperalgesia in the chronic constriction injury model of neuropathic pain //Pain. – 2003. – Т. 104. – №. 1. – С. 229-239.
-
Reinold H. et al. Spinal inflammatory hyperalgesia is mediated by prostaglandin E receptors of the EP2 subtype //Journal of Clinical Investigation. – 2005. – Т. 115. – №. 3. – С. 673.
-
Rexed B. A cytoarchitectonic atlas of the spinal coed in the cat //Journal of comparative neurology. – 1954. – Т. 100. – №. 2. – С. 297-379.
-
Russo C. M., Brose W. G. Chronic pain //Annual review of medicine. – 1998. – Т. 49. – №. 1. – С. 123-133.
-
Rustøen T. et al. Prevalence and characteristics of chronic pain in the general Norwegian population //European Journal of Pain. – 2004. – Т. 8. – №. 6. – С. 555-565.
-
Samad T. A. et al. Interleukin-1beta-mediated induction of Cox-2 in the CNS contributes to inflammatory pain hypersensitivity //Nature. – 2001. – Т. 410. – №. 6827. – С. 471.
-
Schaible H. G., Vanegas H. How do we manage chronic pain? //Best Practice & Research Clinical Rheumatology. – 2000. – Т. 14. – №. 4. – С. 797-811.
-
Scholz J., Woolf C. J. Can we conquer pain? //Nature neuroscience. – 2002. – Т. 5. – №. 11s. – С. 1062.
-
Suen P. et al. NMDA receptor subunits in the postsynaptic density of rat brain: expression and phosphorylation by endogenous protein kinases //Molecular brain research. – 1998. – Т. 59. – №. 2. – С. 215-228.
-
Swett J. E., Woolf C. J. The somatotopic organization of primary afferent terminals in the superficial laminae of the dorsal horn of the rat spinal cord //Journal of Comparative Neurology. – 1985. – Т. 231. – №. 1. – С. 66-77.
-
van Tulder M. W., Koes B., Malmivaara A. Outcome of non-invasive treatment modalities on back pain: an evidence-based review //European Spine Journal. – 2006. – Т. 15. – №. 1. – С. S64-S81.
-
van Tulder M. W. et al. Outcome of invasive treatment modalities on back pain and sciatica: an evidence-based review //European Spine Journal. – 2006. – Т. 15. – №. 1. – С. S82-S92.
-
Watkins L. R., Milligan E. D., Maier S. F. Glial activation: a driving force for pathological pain //Trends in neurosciences. – 2001. – Т. 24. – №. 8. – С. 450-455.
-
Rodriguez R. F. et al. Codeine/acetaminophen and hydrocodone/acetaminophen combination tablets for the management of chronic cancer pain in adults: a 23-day, prospective, double-blind, randomized, parallel-group study //Clinical therapeutics. – 2007. – Т. 29. – №. 4. – С. 581-587..
-
Wilson K. G. et al. Major depression and insomnia in chronic pain //The Clinical journal of pain. – 2002. – Т. 18. – №. 2. – С. 77-83.
-
Woolf C. J. Somatic pain--pathogenesis and prevention //British Journal of Anaesthesia. – 1995. – Т. 75. – №. 2. – С. 169-176.
-
Woolf C. J. Pain: moving from symptom control toward mechanism-specific pharmacologic management //Annals of internal medicine. – 2004. – Т. 140. – №. 6. – С. 441-451.
-
Woolf C. J. Central sensitizationuncovering the relation between pain and plasticity //Anesthesiology: The Journal of the American Society of Anesthesiologists. – 2007. – Т. 106. – №. 4. – С. 864-867.
-
Woolf C. J., Costigan M. Transcriptional and posttranslational plasticity and the generation of inflammatory pain //Proceedings of the National Academy of Sciences. – 1999. – Т. 96. – №. 14. – С. 7723-7730.
-
Woolf C. J., Decosterd I. Implications of recent advances in the understanding of pain pathophysiology for the assessment of pain in patients //Pain. – 1999. – Т. 82. – С. S141-S147.
-
Woolf C. J., Fitzgerald M. Somatotopic organization of cutaneous afferent terminals and dorsal horn neuronal receptive fields in the superficial and deep laminae of the rat lumbar spinal cord //Journal of Comparative Neurology. – 1986. – Т. 251. – №. 4. – С. 517-531.
-
Woolf C. J., Mannion R. J. Neuropathic pain: aetiology, symptoms, mechanisms, and management //The lancet. – 1999. – Т. 353. – №. 9168. – С. 1959-1964.
-
Woolf C. J., Salter M. W. Neuronal plasticity: increasing the gain in pain //science. – 2000. – Т. 288. – №. 5472. – С. 1765-1768.
-
Yu X. M. et al. NMDA channel regulation by channel-associated protein tyrosine kinase Src //Science. – 1997. – Т. 275. – №. 5300. – С. 674-678.
-
Zeilhofer H. U. The glycinergic control of spinal pain processing //Cellular and Molecular Life Sciences. – 2005. – Т. 62. – №. 18. – С. 2027-2035.
-
Zeilhofer H. U., Brune K. Analgesic strategies beyond the inhibition of cyclooxygenases //Trends in pharmacological sciences. – 2006. – Т. 27. – №. 9. – С. 467-474.
-
Zimmermann M. Pathobiology of neuropathic pain //European journal of pharmacology. – 2001. – Т. 429. – №. 1. – С. 23-37.
-
Zirrgiebel U. et al. Characterization of TrkB receptor‐mediated signaling pathways in rat cerebellar granule neurons: Involvement of protein kinase C in neuronal survival //Journal of neurochemistry. – 1995. – Т. 65. – №. 5. – С. 2241-2250.
