
Биологические часы
Одной из самых полезных особенностей организма — возможность предсказывать предстоящие изменения окружающей среды. Это позволяет организму подготовиться к жизненно важным изменениям, которые в противном случае оказались бы неожиданными. Бактериям выгоднее отложить репликацию ДНК до темного времени суток, когда шанс УФ-повреждения резко снижается; мухе — найти прохладное место до того, как полуденное солнце иссушит ее; подъем за несколько минут до раздражающего звона Вашего будильника, вероятно, повысит настроение. Фактически, большинство созданий на нашей планете обладает способностью предвидеть приближающиеся события благодаря их биологическим часам. Эти хронометры настроены на постоянные внешние события — в основном, ежедневные колебания света и температуры.
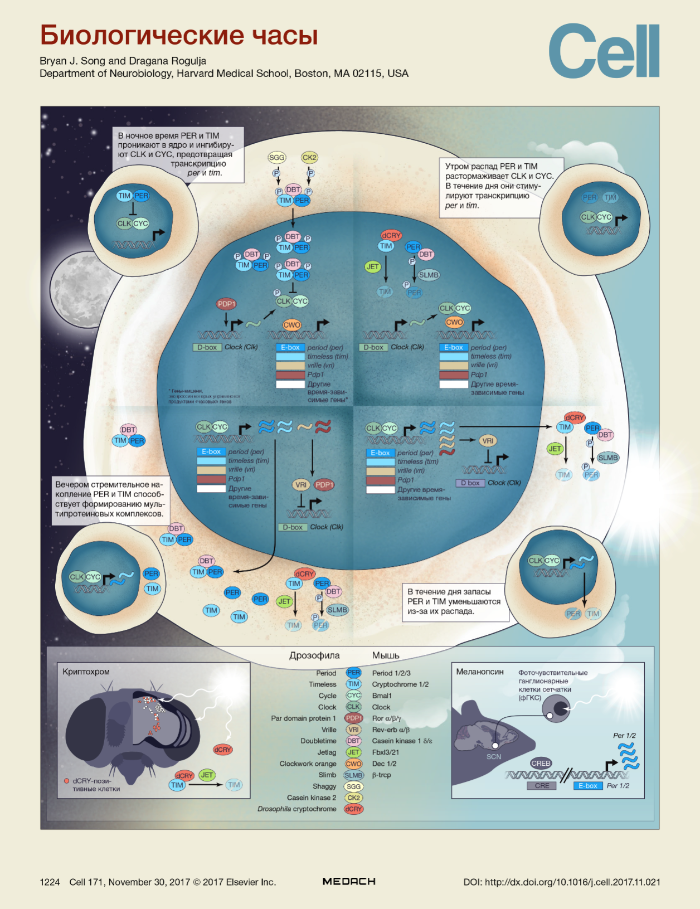
У животных биологические часы «тикают» во всех клетках организма и регулируются «главным часовщиком» в специализированных нейронах головного мозга. В основе циркадных ритмов лежат молекулярные механизмы, впервые открытые в 1971 году, когда Конопка и Бензер сообщили о том, что различные точечные мутации в единичном гене (который позже был клонирован и назван period) могут сдвигать вперед или назад во времени, а также изменять случайным образом регламент повседневных привычек у плодовых мух. Спустя десятилетия исследований мы узнали многое о различных компонентах этих часов, в основе которых лежит транскрипционная/трансляционная цепь отрицательной обратной связи, которая обеспечивает различные клеточные состояния на протяжении дня. Это осуществляется благодаря изменению количества и локализации ключевых мРНК, белков и их комплексов.
Основные компоненты транскрипционной/трансляционной цепи обратной связи
Несмотря на некоторые важные изменения, биологические часы на протяжении всей эволюции остаются крайне простыми как по составу, так и по регуляции, что дает возможность полностью их описать, опираясь на данные о мухах. У дрозофилы существует четыре основных компонента цепи обратной связи — Clock (CLK), Cycle (CYC), Period (PER) и Timeless (TIM). Потеря любого из них приводит к сбою ритмов. У млекопитающих эквивалентом TIM являются криптохромы (CRY), а эквивалентом CYC является BMAL1, при этом выделены три гена per и два гена cry.
В общих чертах регуляция происходит следующим образом: CLK и CYC запускают экспрессию per и tim. PER и TIM, в свою очередь, ингибируют транскрипционную активность CLK и CYC. Таким образом, PER и TIM ингибируют свою собственную экспрессию, и подобное развитие событий приводит к постепенной смене состояний, которая может беспрерывно считываться для определения времени суток.
Повышение концентрации PER и TIM (позитивное плечо обратной связи)
Утром гетеродимеры CLK-CYC начинают связывать последовательности ДНК в регуляторных участках их таргетных генов, называемых Е-боксы, среди которых есть per и tim. В течение дня экспрессия per и tim увеличивается. Накоплению этих белков препятствует фосфорилирование PER киназой Doubletime (DBT; казеинкиназа 1 [CK1] у млекопитающих), которая помечает их для проведения дальнейшего расщепления. У мух накопление TIM замедляется из-за света, попадающего напрямую через тонкую кутикулу и активирующего криптохром дрозофил (dCRY) — фоточувствительный белок, экспрессируемый в специальных «клетках-часовщиках». Связывая TIM, dCRY отмечает его для последующего убиквитинирования с помощью SCF-убиквинтинлигаз, одним из компонентов которых является белок Jetlag (JET) из F-бокса. Динамика накопления PER и CRY у млекопитающих также регулируется через распад, опосредованный SCF-убиквитинлигазами.
Ингибирование активности CLK-CYC (отрицательное плечо цепи обратной связи)
При накоплении PER и TIM в цитоплазме их способность избегать расщепления возрастает. У мух это сильнее всего проявляется в темноте, когда свет больше не может привести к разрушению TIM. В результате формируются комплексы PER-TIM-DBT, в которых PER и TIM более стабильны. В связанном виде TIM фосфорилируется Shaggy, а PER — казеинкиназой 2. Это фосфорилирование регулирует транслокацию комплекса в ядро (реакции, регулирующие проницаемость ядер млекопитающих, до сих пор неизвестны). В ядре PER-TIM-DBT предотвращает связывание CLK-CYC с E-боксами путем взаимодействия с его гетеродимерами и фосфорилирования CLK. Это подавляет экспрессию «часовых» генов, включая per и tim. У млекопитающих комплексы PER-CRY-CK1 не предотвращают связывание CLK-BMAL1 с E-боксами, но привлекают транскрипционные комплексы-репрессоры. Следует отметить, что у млекопитающих комплексы PER-CRY были обнаружены и в цитоплазме, и в ядре.
Ежедневный сброс часов на свету
Поздним вечером устанавливается высокий уровень PER и TIM, который предотвращает дальнейшую транскрипцию per и tim. С наступлением утра свет нарушает ответную петлю обратной связи путем стимуляции транскрипции per и tim, но у мух и млекопитающих этот процесс происходит по-разному. У мух активируемый светом CRY напрямую стимулирует распад TIM. У млекопитающих свет активирует фоточувствительные ганглионарные клетки сетчатки (фГКС), которые экспрессируют фотопигмент меланопсин и запускают сигнал о светлом времени суток по отдельным от классического проводящего пути зрительного анализатора путям. От фГКС сигнал идет напрямую к «часовым» нейронам в супрахиазмальное ядро гипоталамуса, управляющее суточным ритмом, запуская транскрипцию per1 и per2.
Вторичная петля
Экспрессия clk ритмически регулируется посредством вторичной петли, состоящей из CLK-CYC и их прямых мишеней Vrille (VRI) и PDP1. Синтезированные в разное время, VRI и PDP1 могут оказывать противоположное воздействие на экспрессию clk (негативное и позитивное соответственно). Вопреки колебаниям концентрации соответствующей мРНК, концентрация белка CLK значительно не изменяется на протяжении дня. Однако синтез нового CLK утром может сыграть свою роль в замещении инактивированного гиперфосфорилированного CLK, подлежащего расщеплению. У млекопитающих ритмически экспрессируется bmal1, а не clk; ROR и REV-ERB служат эквивалентами VRI и PDP1.
Результат
Авторы описали основные механизмы, запускающие биологические часы. Биологические часы контролируют физиологические процессы, которые должны происходить в определенное время дня. Режим сна и бодрствования является примером результата работы биологических часов. Но и такие процессы, как поддержание температуры тела, пищеварение и деление наших клеток регулируются биологическими часами. Последние исследования говорят о том, что концентрации от 5 % до 20 % различных транскриптов у млекопитающих меняются в зависимости от времени суток. Это значит, что циркадные ритмы влияют на множество (если не на большинство) биологических процессов.
