
CD-ландшафт клеток

Изучая объект, мы пытаемся его охарактеризовать, сравнить с уже известными и найти ему место на полке нашего сознания. Наука и жизнь немыслимы без классификации. Системы рождаются, устаревают, приходят в забвение… Или же успешно трансформируются и развиваются, превращаясь в настоящие звезды. Итак, встречайте — неподражаемые кластеры дифференцировки. У армии их фанатов нет громоздких СD-ROM, но в закладках браузера CD Maps [1, 2].
Кластер дифференцировки (cluster of differentiation, cluster designation, CD) — это маркер, который идентифицирует конкретный паттерн клеточной дифференцировки, выявляемый специфическим моноклональным антителом [3].
Номенклатура CD завоевала официальный статус: она принята научным сообществом и одобрена Международным союзом иммунологических обществ и Всемирной организацией здравоохранения.
Рожденные гибридомной революцией
Возникновению системы CD способствовало получение моноклональных антител с уникальной специфичностью (Георг Келер, Цезарь Мильштейн, 1975 год) [4]. Это стало возможным благодаря разработке метода гибридом, воплощающего мечту «приставить губы Никанора Ивановича к носу Ивана Кузьмича». Соматический гибрид нормальной антителообразующей и опухолевой клетки (гибридома) передает своим потомкам как бессмертие злокачественно трансформируемой клетки, так и возможность синтезировать антитела. Белки имеют специальный узор из опознавательных знаков — детерминантных групп, каждая из которых представлена несколькими остатками аминокислот или сахаров. То есть один белок имеет несколько различных детерминант и, следовательно, широкий спектр антител, с которыми возможно образование связи. Узнавание молекулы антителом подразумевает образование с ней значительно более прочной связи по сравнению с другими молекулами. Крепость «уз» в данном случае измеряется сродством или константой диссоциации. Для многих исследований требуются структуры с более четкими характеристиками. Моноклональные антитела нацелены на одну конкретную детерминанту, а их физико-химическая однородность превращает их в высокочувствительные реагенты [5].
Открывшиеся перспективы поражали воображение, и радостные иммунологи генерировали все большее количество антител. Однако новой технологии отчаянно не хватало упорядоченности. Иногда полученные в разных лабораториях разноименные структуры фактически распознавали одни и те же паттерны. Это привело к хаотичному называнию молекул — Вавилонской башне терминологии [6].
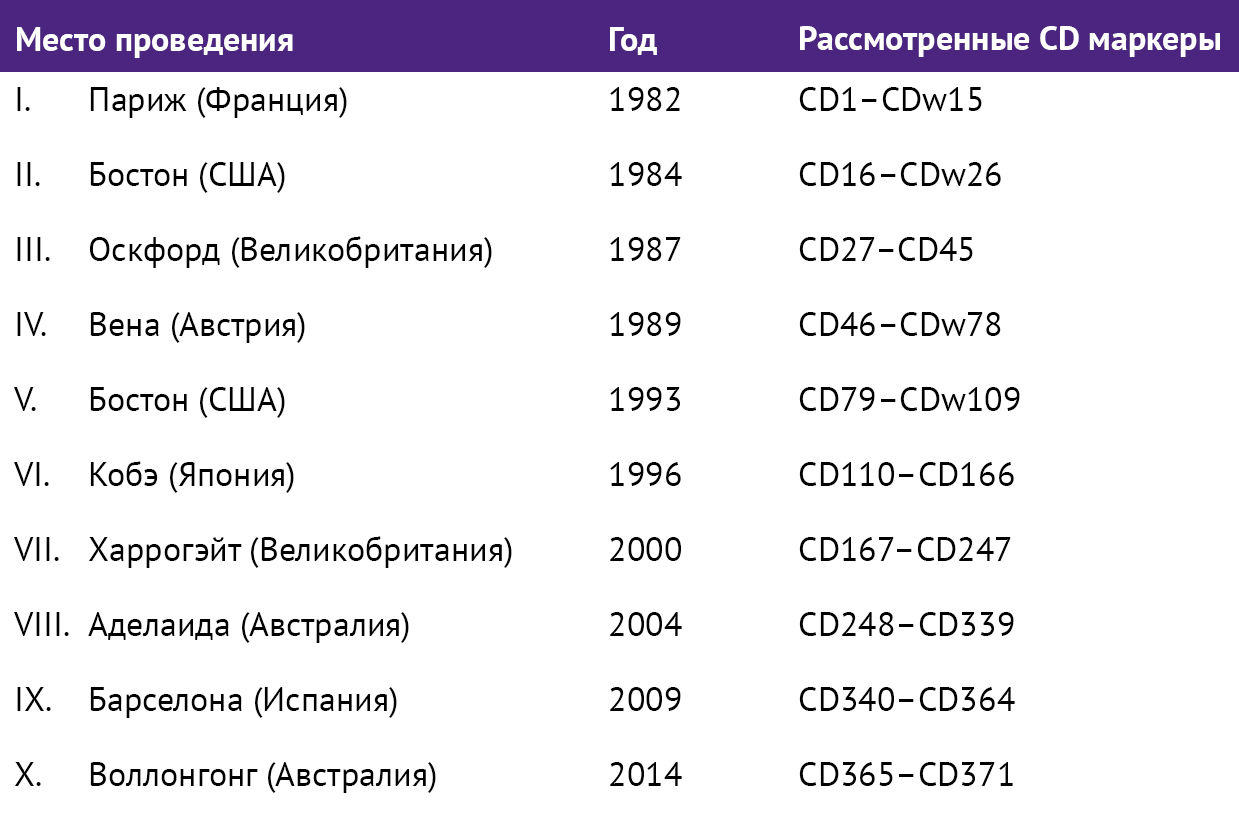
Для ликвидации все нарастающей путаницы в 1982 году Ален Бернард и Лоуренс Бумселл организовали в Париже I Международное рабочее совещание по дифференцировочным антигенам лейкоцитов человека (the I International Workshop of Нuman Leukocyte Differentiation Antigens, HLDA). 55 коллективов ученых из 14 стран согласованно работали по единому протоколу. В итоге удалось объединить исследованные на тот момент антигены в 15 кластеров, обозначенных буквами CD [7].
Мультилабораторный слепой анализ антител обеспечил независимую проверку специфичности молекул и послужил основой для уверенного использования этих реагентов в фундаментальных исследованиях и клинической практике. Сложные коммуникации клеток иммунной системы и невозможность рассматривать ее изолированно привели к расширению объектов исследований экспертов HLDA. На сегодняшний день, помимо классического анализа лейкоцитов, в качестве объектов рассматриваются и другие типы клеток: гемопоэтические стволовые, кроветворные клетки-предшественницы, тромбоциты, дендритные и эндотелиальные клетки. В настоящее время проведение рабочих совещаний HLDA осуществляет неправительственная организация Молекулы дифференцировки клеток человека (Human Cell Differentiation Molecules, HCDM) со штаб-квартирой в Барселоне (Испания). Актуальный список маркеров включает 371 CD [8].
Таблица 1 | Рабочие совещания HLDA I–X (1982–2014) [8]
Термин CD сбросил чешую и оброс перьями, то есть успешно эволюционировал. Строгое определение СD как поверхностных белков лейкоцитов утратило свою актуальность. Не все CD — белки, не все поверхностные, не все встречаются на лейкоцитах. Научный прогресс вынуждает отказываться от категоричных определений фундаментальных свойств, чтобы избежать необходимости постоянных уточнений и абсурдных ситуаций, когда исключений больше, чем соответствий правилу. Рационально вводить четкие критерии, основанные на воспроизводимых параметрах.
Для признания нового CD требуется представить на суд инквизиторов HCDM свидетелей — моноклональные антитела из независимых лабораторий с идентичным характером реактивности, которые к тому же опознают одну и ту же молекулу. Протокол заседания строго контролируется. Основные лаборатории-участники тестируют реактивность антител с несколькими типами клеток, используя многоцветную проточную цитометрию. В других лабораториях проводят проверку специфической реактивности с использованием методов иммунобиохимии (иммунопреципитация, вестерн-блоттинг) и иммуногистохимии. Моноклональные антитела должны специфически распознавать как антиген в трансфицированных клетках, так и его эндогенный аналог в первичных клеточных линиях [3].
- Проточная цитометрия — метод исследования дисперсных сред в режиме поштучного анализа элементов дисперсной фазы по сигналам светорассеивания (прямое светорассеивание — для определения относительного размера клеток или частиц; боковое светорассеивание — для оценки неоднородности внутриклеточного содержимого клетки, например, размеров ядра и гранулярности цитоплазмы) и флуоресценции (изучение клеточных маркеров с помощью меченных флюорохромными красителями антител к поверхностным и внутриклеточным компонентам клеток) [9].
- Иммунопреципитация — способ, с помощью которого можно выделить из смеси и осадить («precipitate») искомую молекулу за счет образования комплекса антиген-антитело.
- Вестерн-блоттинг или иммуноблотинг («blotting» — промокание) — аналитический метод, основанный на комбинации гель-электрофореза (проведение электрофоретического разделения белков) и иммунохимической реакции «антиген/антитело-исследуемый белок».
Иммуногистохимия — метод выявления специфических антигенов в тканях в результате распознавания соответствующим антителом с последующим анализом микропрепаратов на светооптическом уровне [10]. - Трансфекция — метод генной инженерии, заключающийся в изменении фенотипа путем введения в клетку (эукариотическую) чужеродной нуклеиновой кислоты без использования вирусов. Вирусная «доставка» нуклеиновой кислоты называется трансдукцией [11].
Вопрос, что было раньше, не решен для пары курица и яйцо, но определен для моноклонального антитела и идентифицируемой им молекулярной частицы. Изначально именно моноклональное антитело использовалось для характеристики своей мишени. Например, CD2-моноклональные антитела представляют собой реагенты, которые реагируют с трансмембранным гликопротеином с молекулярной массой 50 кДа, экспрессируемым в покоящихся Т-клетках. В настоящее время клеточные структуры сначала идентифицируются с помощью методов молекулярной генетики или протеомики, а затем уже моделируются специфические антитела [12].
Строчная буква «w» («workshop»), предшествующая обозначению номера, используется для еще не утвержденных кандидатов. Например, молекула все еще в листе ожидания, т. к., по данным совещаний HLDA, для нее подтверждено только одно специфическое антитело.
Анализ w-клеймированных маркеров, рассмотренных еще в начале деятельности HLDA, выявил их принадлежность к кластерам моноклональных антител, распознающих углеводные эпитопы, которые после надлежащей биохимической идентификации получили свой собственный «чистый» номер CD. Например, антиген Томсена-Фриденрайха (TF или T) открыт случайно при изучении групп крови (обнаруживался на контаминированных эритроцитах). Структурно TF — это универсальная первичная (коровая, кор-1) последовательность O-гликанов, то есть углеводный эпитоп. Присутствуя практически на всех мембранных гликопротеинах муцинового типа, он остается иммунологически замаскированным из-за удлинения углеводной цепи. Но в злокачественно трансформированных клетках происходит его демаскировка, вероятнее всего, в результате нарушения (обрыва) гликозилирования. TF характеризуется как онкофетальный углеводный эпитоп, имеет номер CD176. Он определяется примерно в 90 % опухолей преимущественно эпителиального строения (толстая кишка, молочная железа, мочевой пузырь, простата, печень, яичники и желудок), при этом уровень экспрессии вариабелен [13].
Прописные буквы после номера CD обозначают сплайсированный вариант внеклеточного домена молекулы клеточной поверхности. Например, в результате альтернативного сплайсинга гена Ptprc (Protein tyrosine-phosphatase, receptor type), кодирующего трансмембранную тирозиновую протеинфосфатазу CD45, образуются варианты с различными характеристиками. Наивные Т-лимфоциты экспрессируют преимущественно высокомолекулярные изоформы СD45 (CD45RA), имеют высокую фосфатазную активность и поддерживают Т-клеточный рецептор в премированном состоянии для распознавания антигена. Активированные Т-клетки экспрессируют короткий сплайс-вариант CD45 (CD45RO), с которым связывают более быструю и эффективную активацию, опосредованную антигеном [14].
- Сплайсинг — это процесс созревания молекул, в результате которого из предшественника удаляется внутренняя часть, а края образовавшегося дефекта лигируются за счет образования ковалентной связи. Сплайсингу подвергаются нуклеиновые кислоты и белки. Вырезание разных участков в транскриптах формирует альтернативный сплайсинг, в результате которого с одного гена считываются разные белки [15].
Строчная буква после номера CD обозначает несколько молекул, которые имеют общую цепь. Например, CD1a, CD1b, CD1c, CD1d, CD1e имеют цепь β2-микроглобулина. Интегрин бета-2 (CD18), связываясь с различными альфа-субъединицами интегрина, образует гетеродимерные комплексы (CD11a, CD11b и CD11c). В других случаях строчные буквы используются для обозначения разных членов одного и того же семейства генов, как в случае с CD66 (CD66a, CD66b, CD66c, CD66d, CD66e и CD66f).
В случае углеводных структур CD строчная буква указывает на модификацию углеводной последовательности. Например, CD65s трактуется как сиалилированный CD65; CD60b и CD60c означают 9-O- и 7-О-ацетилированный ганглиозид GD3 [3].
Символ «+» или «–» указывает на наличие или отсутствие экспрессии данной молекулы в определенном типе клеток. В некоторых случаях экспрессия уточняется количественно: «высокий» или «низкий» уровень.
Поверхность цитоплазматической мембраны — важный участник всех видов клеточной коммуникации, фактически, она осуществляет ведение внешней политики клетки. Как настоящий дипломат (студент МГИМО в случае наивных лимфоцитов, или опытный спец — клетка памяти), клеточная поверхность реагирует на потенциально опасные изменения окружающей среды, опосредует клеточную адгезию и коммуникацию между клетками (как внутри иммунной системы, так и со стромой). В состав министерства внутренних дел входят рецепторы, транспортеры, каналы, белки адгезии клеток, ферменты. Поверхностно экспрессируемые белки получают мощную «государственную» поддержку — около 26 % человеческих генов кодируют примерно 5500 трансмембранных белков [16]. Согласно оценкам in silico, 2886 таких белков фактически экспрессируются на наружной клеточной мембране, то есть непосредственно на клеточной поверхности [17]. Экспериментальные данные представлены для 1492 белков разных тканей [18]. 1015 исследованных белков обнаруживаются в одном или нескольких типах иммунных клеток, в лимфоидной ткани [19].
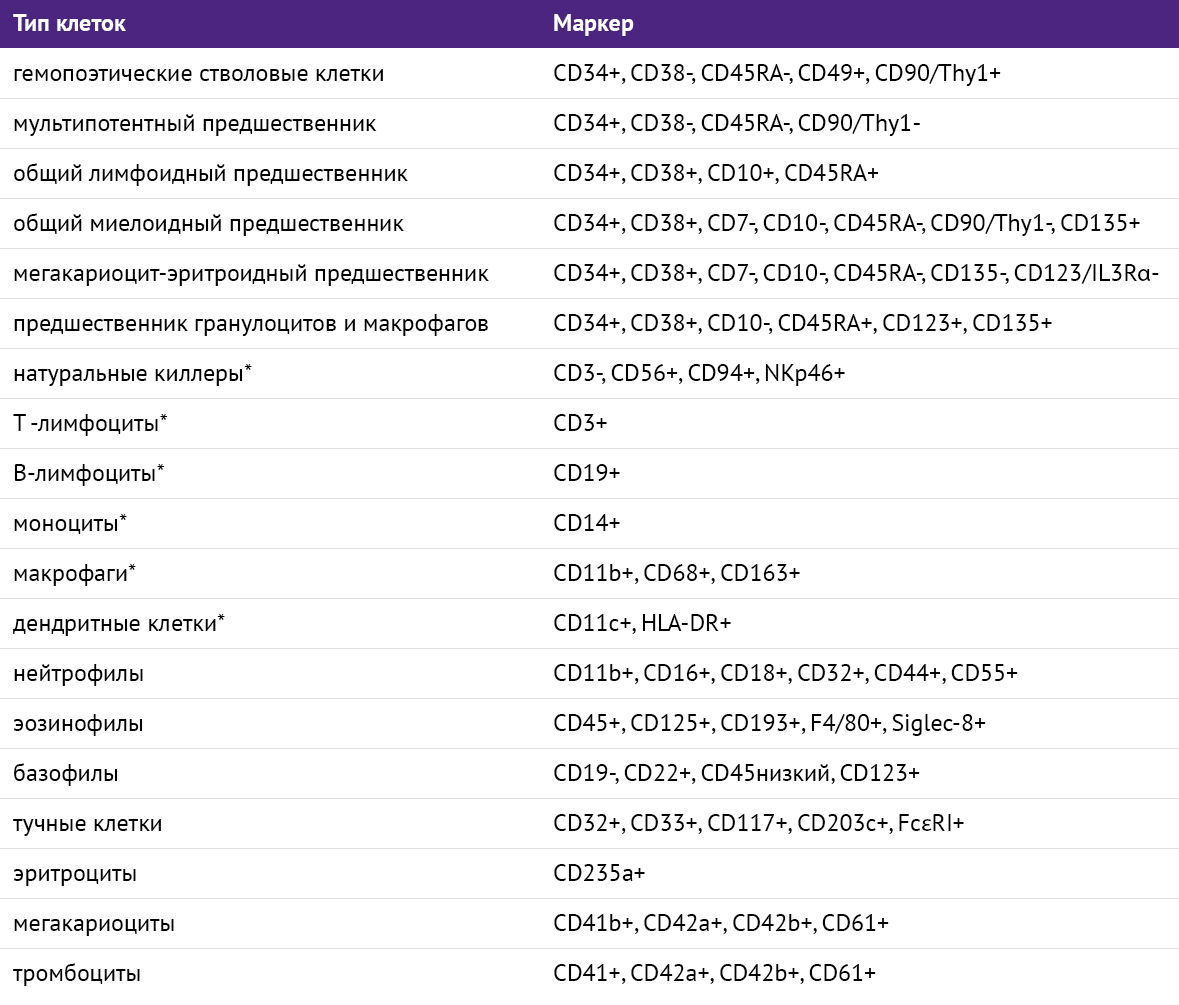
Ландшафт клеточной поверхности все еще загадочен. Заявленные внушительные цифры требуют тщательного анализа, уточнения и унификации информации. На фоне хаотичного разнообразия разношерстных молекул выгодно выделяются CD-маркеры (исторически определенные как белки клеточной поверхности и представляющие большинство из существующих маркеров). Длительная история исследований и ранний акцент на стандартизации методов превращает их в достаточно удобный инструмент, который позволит набросать эскиз карты клеточной поверхности и далее дополнять его (табл. 2) [20,21].
Таблица 2 | Основные дифференцировочные маркеры клеток крови [20,21]
При изучении белков используются методы сравнительной геномики, базирующиеся на следующем представлении: биомолекулы, имеющие значительное сходство на уровне последовательностей, имеют сходные структуры и функции. При этом возможно обнаружить сходство на различных структурных уровнях (гомологию), что указывает на их эволюционную взаимосвязь. Гомологичные белки образуются в результате различных событий. Например, при расхождении в ходе видообразования сходные гены обнаруживаются у разных видов. Такие белки (и кодирующие их гены) являются ортологами. У одного вида ген может дуплицироваться (то есть образовать две копии), далее каждая копия развивается по-своему — так возникают паралоги [23].
ФИО клетки прочно ассоциируется с номерами CD (рис. 1) [21–22,24–26]. Анализ экспрессионного статуса молекул CD в рамках проведения иммунофенотипирования является фундаментальным компонентом диагностики, классификации и мониторинга гемобластозов, а также аутоиммунных заболеваний и иммунодефицитов [27,28]. Морфологическое исследование на светооптическом уровне не позволяет отличить различные типы лимфоцитов, отследить степень их дифференцировки.
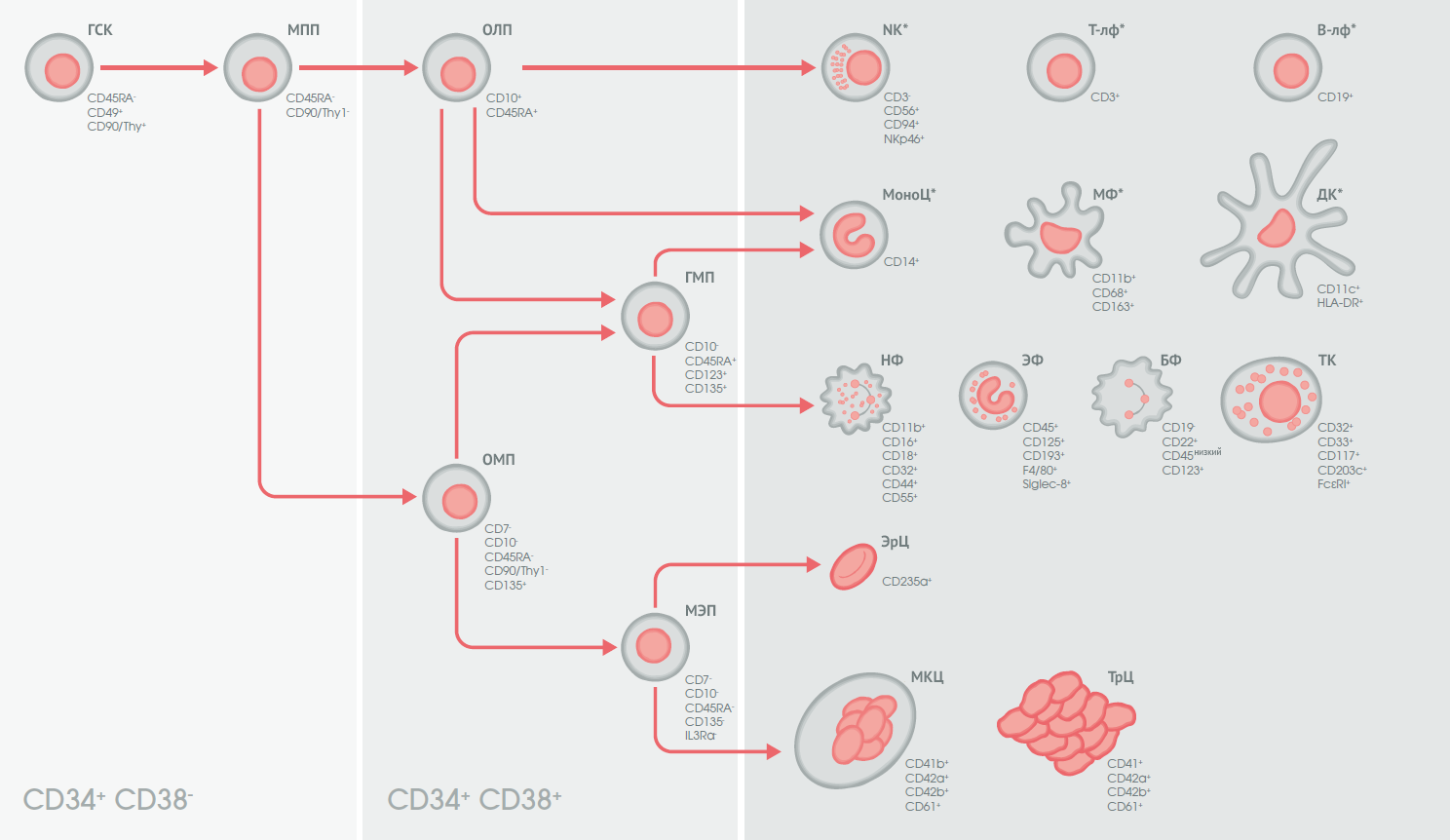
ГСК — гемопоэтические стволовые клетки;
МПП — мультипотентный предшественник;
ОЛП — общий лимфоидный предшественник;
ОМП — общий миелоидный предшественник;
МЭП — мегакариоцит-эритроидный предшественник;
ГМП — предшественник гранулоцитов и макрофагов;
NK* — NK-клетки (натуральные киллеры)*;
Т-лф* — T -лимфоциты*;
В-лф* — B-лимфоциты*;
МоноЦ*— мoноциты*;
МФ*— мaкрофаги*;
ДК* — дендритные клетки*;
НФ — нейтрофилы;
ЭФ — эозинофилы;
БФ — базофилы;
ТК — тучные клетки;
ЭрЦ — эритроциты;
МКЦ — мегакариоциты;
ТрЦ — тромбоциты.
CD123/IL3Rα — альфа-субъединица рецептора интерлейкина 3;
NKp46 — рецептор естественной цитотоксичности 1 (NCR1), пан-NK-маркер;
HLA-DR (Human Leucocyte Antigen — антиген лейкоцитов человека) — антиген человеческого главного комплекса гистосовместимости класса II, маркер иммуногенной активации;
F4/80 — маркер популяций мышиных макрофагов. Человеческий ортолог этого белка называется EMR1: модуль, подобный человеческому эпидермальному фактору роста (EGF), содержащий рецептор муцинподобного гормона 1, является высокоспецифичным маркером эозинофилов [22];
Siglec-8 — иммуноглобулин-подобный сиало-связывающий лектин 8 (Sialic acid-binding Ig-like lectins 8) [24];
FcεRI — высокоаффинный Fc-эпсилон-рецептор I для IgE
Четкие линии эскиза появляются при хорошо заточенном карандаше. Обострить точность планируется и для CD-маркеров. В наших знаниях о паттернах экспрессии молекул CD все еще остаются значительные пробелы, в том числе из-за неоднородности исследований экспрессии и значительных изменений в технологиях проточной цитометрии за 30 лет проведения семинаров HLDA.
Организация HCDM инициировала проект CD Maps — многопрофильную исследовательскую программу для создания карты клеточной поверхности иммунных клеток человека. Планируется проверить экспрессию уже известных CD маркеров на основных клеточных типах, используя современные возможности лабораторной диагностики (рис. 2) [1]. Точность данных обеспечивается использованием стандартизированных протоколов многоцветной проточной цитометрии. CD Maps должен создать огромную базу данных профилей экспрессии молекул, которая будет представлять собой уникальный ресурс с открытым доступом для фундаментальной, трансляционной и клинической иммунологии.
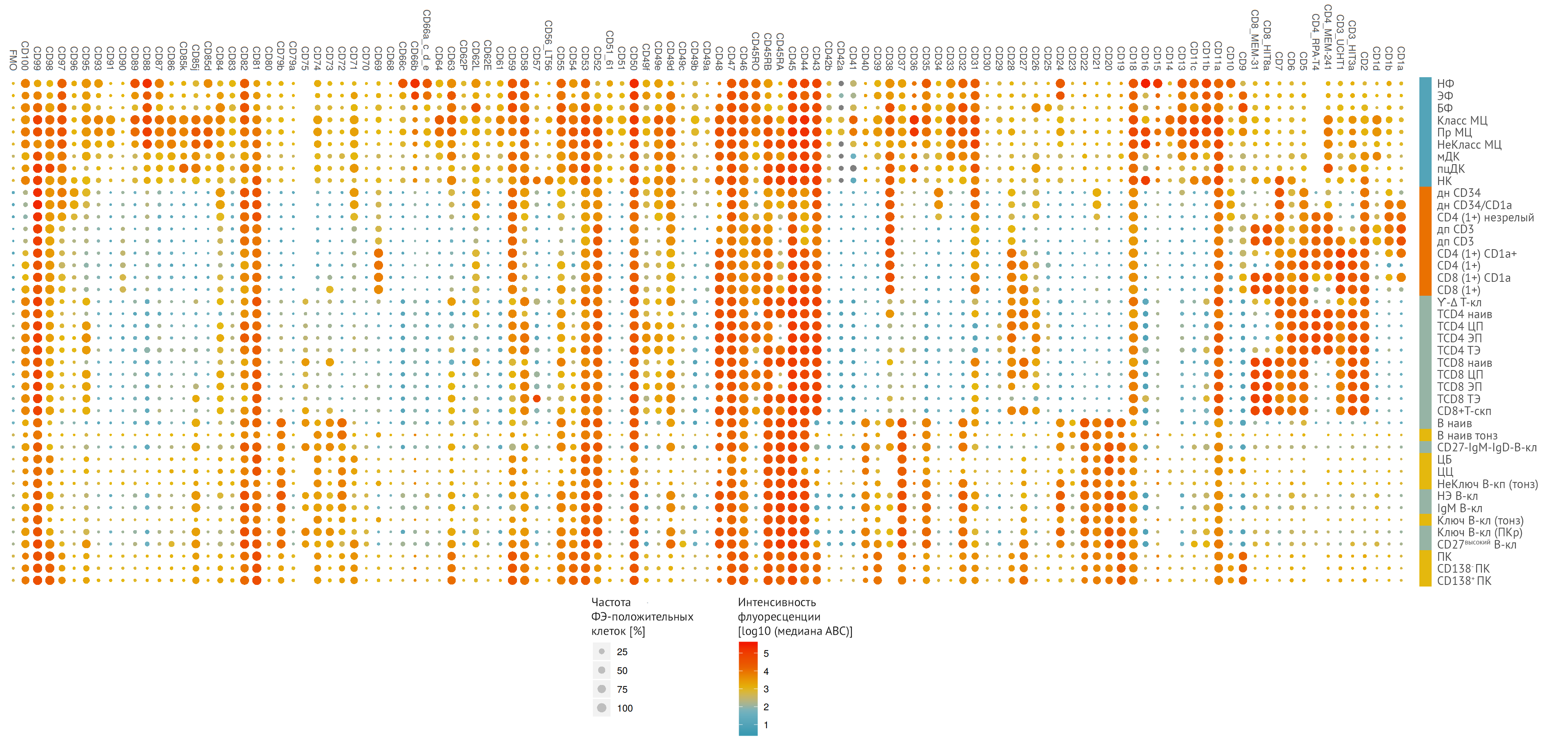
Интенсивность флуоресценции [log10 (медиана ABC (антигенсвязывающая способность)]. Для проведения иерархического кластеризационного анализа экспрессии маркеров CD клеточные популяции лейкоцитов были сформированы в 10 подгрупп:
1. Гранулоциты.
НФ — нейтрофилы; ЭФ — эозинофилы; БФ — базофилы
2. Моноциты.
НеКласс МЦ — неклассические моноциты; Класс МЦ — классические моноциты; Пр МЦ — промежуточные моноциты.
3. Дендритные клетки.
мДК — миелоидные дендритные клетки; пцДК — плазмоцитоидные дендритные клетки.
4. Натуральные киллеры (НК).
5. Тимоциты.
днCD34 — двойной негативный CD34; днCD34/CD1a — двойной негативный CD34/CD1a.
6. CD4+ T-клетки.
CD4(1+) незрелый — незрелый единичный позитивный CD4; CD4 (1+) CD1a+ — единичный позитивный CD4 и позитивный CD1a; CD4 (1+) — единичный позитивный CD4; TCD4 наив — наивные CD4+ T-клетки; TCD4 ЦП — CD4+ T-клетки центральной памяти; TCD4 ЭП — CD4+ T-клетки эффекторной памяти; TCD4 ТЭ — терминальные эффекторные CD4+ T-клетки, реэкспрессирующие CD45RA (TEMRA).
7. CD8+ T-клетки.
CD8 (1+) CD1a — единичный позитивный CD8 и позитивный CD1a; CD8 (1+) — единичный позитивный CD8; TCD8 наив — наивные CD8+ T-клетки; TCD8 ЦП — CD8+ T-клетки центральной памяти; TCD8 ЭП — CD8+ T-клетки эффекторной памяти; TCD8 ТЭ — терминальные эффекторные CD8+ T-клетки, реэкспрессирующие CD45RA (TEMRA); CD8+T-скп — популяция CD8+Т-лимфоцитов–стволовые клетки памяти, отражающая переход наивных Т-клеток в клетки памяти с экспрессией CD27+, CD28+, CD95, а также CD45RА+ и CD45RO+ (CD45RА dim — «тусклый» фенотип) с высокой способностью к пролиферации.
8. Другие Т-клетки.
дпCD3 — двойной позитивный CD3; ϒ-Δ Т-кл — гамма-дельта Т-клетки.
9. В-клетки
B наив — наивные В-клетки; В наив тонз — тонзиллярные наивные В-клетки; CD27-IgM-IgD В-кл — CD27-IgM-IgD В-клетки; ЦБ — центробласты; ЦЦ — центроциты; Ключ и НеКлюч В-кл (тонз) — переключенные и непереключенные В-клетки памяти, полученные из ткани миндалин; НЭ B-клетки — натуральные эффекторные В-клетки; IgM B-кл — B-клетки, экспрессирующие только IgМ; Ключ В-кл (ПКр) — переключенные В-клетки памяти, полученные из периферической крови; СD27высокий В-кл — В-клетки с высоким уровнем экспрессии СD27 (СD27 — это маркер терминальной дифференцировки для В-лимфоцитов, экспрессируется на умеренном уровне в В-клетках памяти и на высоком уровне — в плазматических клетках); ПК — плазматические клетки; CD138- ПК — CD138-негативные плазматические клетки; CD138+ ПК — CD138-позитивные плазматические клетки.
10. Лимфоциты — без уточнения подтипа.
Уровень экспрессии маркера CD отражен в цветовой кодировке (негативная — синий, позитивная — красный цвет), размер точек соответствует частоте выявленных используемым флуорохромом фикоэритрином (ФЭ-положительных) клеток в процентах, обе переменные являются медианами всех измеренных образцов [1].
Источники:
- Kalina T.e al.CD Maps-Dynamic Profiling of CD1-CD100 Surface Expression on Human Leukocyte and Lymphocyte Subsets. Front Immunol. 2019;10:2434.
- http://bioinformin.cesnet.cz/
- Engel P.et al.CD Nomenclature 2015: Human Leukocyte Differentiation Antigen Workshops as a Driving Force in Immunology. J Immunol.2015;195(10):4555-4563.
- Kohler G, Milstein C. Continuous cultures of fused cells secreting antibody of predefined specificity. Nature.1975; 256: 495–497.
- Posner J., Barrington P., Brier T., Datta-Mannan A. Monoclonal Antibodies: Past, Present and Future. Handb Exp Pharmacol. 2019;260:81-141.
- Baker M. Reproducibility crisis: Blame it on the antibodies. Nature 2015; 521: 274–276.
- Bernard A., Boumsell L. The clusters of differentiation (CD) defined by the First International Workshop on Human Leucocyte Differentiation Antigens. Hum Immunol 1984; 11: 1–10.
- http://www.HCDM.org
- Vembadi A., Menachery A., Qasaimeh M.A. Cell Cytometry: Review and Perspective on Biotechnological Advances. Front Bioeng Biotechnol.2019;7:147.
- Kaufmann S.H.E. Immunology's Coming of Age [published correction appears in Front Immunol. 2019;10:684.
- Uludag H., Ubeda A., Ansari A. At the Intersection of Biomaterials and Gene Therapy: Progress in Non-viral Delivery of Nucleic Acids. Front Bioeng Biotechnol. 2019;7:131.
- Clark G. et al. Nomenclature of CD molecules from the Tenth Human Leucocyte Differentiation Antigen Workshop. Clin Transl Immunology 2016; 5 (1): e57.
- Li F.et al.. Cell surface Thomsen-Friedenreich proteome profiling of metastatic prostate cancer cells reveals potential link with cancer stem cell-like phenotype. Oncotarget. 2017;8(58):98598–98608.
- Rheinländer A., Schraven B , Bommhardt U. CD45 in human physiology and clinical medicine. Immunol Lett. 2018;196:22-32.
- Lewandowska M.A. The missing puzzle piece: splicing mutations. Int J Clin Exp Pathol. 2013;6(12):2675-82.
- Fagerberg L.et al.. Prediction of the human membrane proteome. Proteomics. 2010;10:1141–9.
- Bausch-Fluck D. et al. The in silico human surfaceome. Proc Natl Acad Sci USA. 2018;115:E10988–97.
- Bausch-Fluck D. et al. A mass spectrometric-derived cell surface protein atlas. PLoS ONE. 2015;10:e0121314.
- Diaz-Ramos M.C., Engel P., Bastos R. Towards a comprehensive human cell-surface immunome database. Immunol Lett. 2011;134:183–7.
- National Center for Biotechnology Information (NCBI) [database on the Internet]. U.S. National Library of Medicine at the National Institutes of Health, 2019.
- https://www.abcam.com/
- Legrand F. et al. The eosinophil surface receptor epidermal growth factor-like module containing mucin-like hormone receptor 1 (EMR1): a novel therapeutic target for eosinophilic disorders. J Allergy Clin Immunol. 2014;133(5):1439-47, 1447.e1-8.
- Kustatscher G. et al.Co-regulation map of the human proteome enables identification of protein functions. Nat Biotechnol. 2019;37(11):1361-1371.
- Legrand F. et al. Sialic acid-binding immunoglobulin-like lectin (Siglec) 8 in patients with eosinophilic disorders: Receptor expression and targeting using chimeric antibodies. J Allergy Clin Immunol. 2019;143(6):2227-2237.e10.
- Kneissl S. et al. CD19 and CD20 targeted vectors induce minimal activation of resting B lymphocytes. PLoS One. 2013;8(11):e79047.
- Walker J.A., Smith K.G. CD22: an inhibitory enigma. Immunology. 2008;123(3):314–325.
- Iwai Y., Hamanishi J., Chamoto K., Honjo T. Cancer immunotherapies targeting the PD-1 signaling pathway. J Biomed Sci. 2017; 24: 26.
- Rosman Z., Shoenfeld Y., Zandman-Goddard G. Biologic therapy for autoimmune diseases: an update. BMC Med.2013; 11: 88.
