
Шаг вперед в изучении мейотического деления клетки

Ключевой процесс полового размножения — обмен ДНК между парами хромосом. Выясняется, что один из этапов этого процесса — осуществление разрывов ДНК ферментом Spo11 — зависит от конденсации белков в жидкие капли.
Наши клетки имеют по две копии каждой хромосомы, которые известны как гомологичные хромосомы или гомологи, каждая из которых наследуется от отдельного родителя. Половое размножение невозможно без формирования половых клеток, в каждой из которых содержится только одна копия каждой хромосомы; слияние двух половых клеток в ходе оплодотворения восстанавливает исходное число хромосом для следующего поколения. Половые клетки образуются в ходе особого способа клеточного деления, называемого мейозом, на раннем этапе которого происходит разделение гомологов по отдельным дочерним клеткам. Ошибки разделения хромосом в мейозе могут привести к тому, что в образующихся половых клетках окажется слишком много или слишком мало хромосом. Такая патология называется анеуплоидией и лежит в основе развития, например, синдрома Дауна, а также является основной причиной выкидышей. В своей статье в «Nature» Claeys Bouuaert с соавт. [1] подчеркивают ключевую роль процесса, называемого разделение фаз на границе жидкостей, и лежащего в основе этого ключевого биологического пути.
Точное разделение хромосом в мейозе требует того, чтобы каждая хромосома вначале была идентифицирована и физически связана со своим партнером-гомологом. Эти шаги зависят от пути репарации ДНК, называемого гомологичной рекомбинацией, который начинается с запрограммированного разрыва ДНК в нескольких случайно выбранных сайтах вдоль каждой хромосомы. Разорванные концы ДНК «ищут» схожие последовательности на других хромосомах, в конечном итоге определяя партнера-гомолога и устанавливая физические связи, называемые кроссоверами. Кроссоверы также позволяют обмениваться генетической информацией между гомологичными хромосомами, обеспечивая генетическую изменчивость между родителями и потомством.
Молекулярные механизмы, контролирующие гомологичную рекомбинацию в мейозе, изучаются более двух десятилетий с момента идентификации десяти белков в почкующихся дрожжах Saccharomyces cerevisiae, необходимых для образования разрывов ДНК во время мейоза [2]. Четыре из этих белков составляют ядерный комплекс Spo11 [3, 4], который создает разрывы в ДНК. Три других образуют комплекс MRX, который опосредует этапы процессинга после разрывов [2]. Роли остальных трех белков — Rec114, Mei4 и Mer2, что вместе называются комплексом RMM, — остаются в основном неясными.
Белки комплекса RMM физически связаны друг с другом, и их можно обнаружить в мейотических хромосомах раньше, чем большинство других белков. Это указывает на то, что главную роль в вовлечении основного комплекса Spo11 в участки разрывов спирали ДНК играют именно белки RMM [5]. Claeys Bouuaert с соавт. проливают свет на этот процесс. Для начала авторы провели очистку комплекса RMM из S. cerevisiae и обнаружили, что Rec114 и Mei4 образуют субкомплекс, который связывается с Mer2. Затем ученым удалось показать, что очищенные белки RMM могут конденсироваться на ДНК в форме жидких капель, содержащих сотни копий каждого белка.
Способность некоторых белков конденсироваться в жидкие капельки известна как разделение фаз на границе жидкость-жидкость (LLPS), что лежит в основе множества клеточных процессов, таких как организация генома и процессинг РНК, а также множества способов молекулярной сигнализации [6]. Как сообщалось ранее, LLPS играет ключевую роль в мейозе, управляя сборкой синаптонемного комплекса, удерживающего гомологичные хромосомы вместе и участвующего в последних стадиях рекомбинации генетической информации и формирования кроссоверов [7]. Claeys Bouuaert с соавт. обнаружили, что очищенные формы белков RMM образуют жидкие конденсационные структуры на ДНК (суммарное количество вещества не превышает несколько наномолей). Это является свидетельством того, что обнаруженный процесс LLPS — ключ к пониманию биологических функций этих белков. Подтверждая свои выводы, ученые продемонстрировали, что мутация в Mer2 предотвращает образование подобных конденсатов in vitro, а это в значительной степени затрудняет разрыв спирали ДНК в клетках с помощью комплекса Spo11.
В отдельном исследовании [4], опубликованном в этом году, исследовательской группой под руководством Claeys Bouuaert сообщалось о первой удачной очистке и биохимическом описании ядерного комплекса Spo11, что стало важным шагом в понимании молекулярных механизмов мейотических разрывов ДНК. Используя преимущества этого очищенного комплекса в настоящей работе, ученые продемонстрировали, что конденсаты RMM способствуют вовлечению ядерного комплекса Spo11 в структуру ДНК. Мутация в Rec114, нарушающая связывание комплекса RMM с ядерным комплексом Spo11, также препятствует разрывам ДНК при мейозе. Все это доказывает то, что комплекс RMM необходим для вовлечения ядерного комплекса Spo11 в процесс разрывов ДНК.
Все вместе эти сведения раскрывают роль комплекса RMM как ключевого участника рекомбинации генетической информации при мейозе, способного к самосборке (благодаря процессу LLPS) для вовлечения ядерного комплекса Spo11 в процесс разрывов ДНК по всему геному (рис. 1). Высокая эволюционная консервативность белков RMM служит убедительным доказательством того, что этот механизм остался неизменным у большинства размножающихся половым путем организмов.
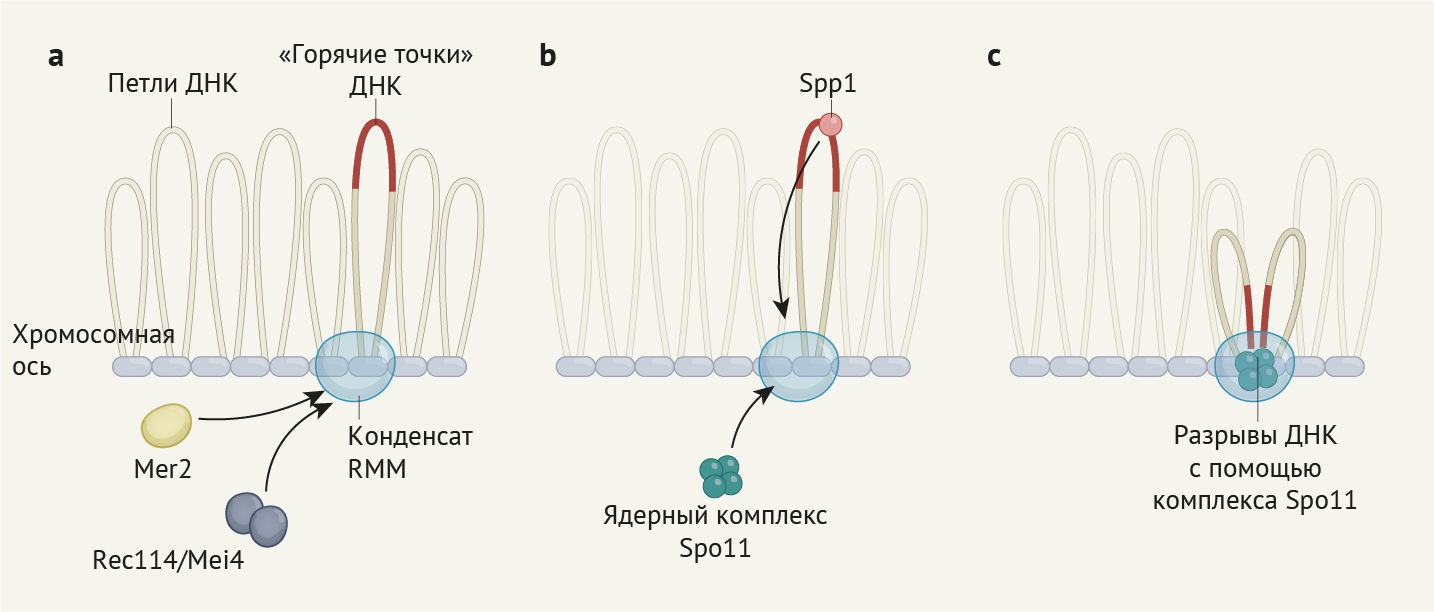
а — На ранних стадиях процесса вокруг богатой белком структуры из хромосом формируются петли ДНК, называемые хромосомной осью. Как сообщают Claeys Bouuaert с соавт. [1], белковый субкомплекс, включающий Rec114 и Mei4, вместе с белком Mer2 конденсируется в похожие на жидкость капельки (т. н. конденсаты RMM) посредством процесса, называемого разделением фаз на границе жидкость-жидкость.
b — Возможно, конденсаты RMM связываются с областями ДНК, называемыми горячими точками, посредством взаимодействия с белком Spp1, а также задействуют в процесс ядерный комплекс Spo11.
c — Spo11 катализирует образование разрывов ДНК — ключевого этапа мейоза.
Работа ученых поднимает несколько вопросов.
- Как разделение фаз может регулировать количество и четкую локализацию разрывов ДНК в геноме во время мейоза? Claeys Bouuaert с соавт. полагают, что конденсация белков RMM в определенных участках хромосомы может приводить к истощению запаса субъединиц RMM в окружающей среде (применительно к данной ситуации — в растворе), что ингибирует образование конденсатов впоследствии и тем самым ограничивает суммарное количество разрывов ДНК, катализируемых в отдельно взятой клетке.
Поскольку разрывы ДНК образуются преимущественно в определенных «горячих точках» в каждой хромосоме, второй вопрос заключается в следующем: - как RMM взаимодействует с другими белками, связанными с мейотическими хромосомами, которые помогают определять расположение «горячих точек»? К этим белкам относятся Hop1, который является частью богатой белком структуры, называемой хромосомной осью, формирующейся в начале мейоза. Hop1 способствует организации хромосом в виде массивов петель ДНК и, вероятно, служит для привлечения RMM к оси путем связывания с Mer28. Также к описываемой группе белков относится Spp1, который распознает молекулярные модификации гистоновых белков, связанных с ДНК в «горячих точках», и может связывать эти последовательности с конденсатами RMM для формирования разрывов [8–10].
И наконец, по-прежнему неизвестно, регулируют ли конденсаты RMM более поздние стадии генетической рекомбинации в мейозе после разрыва ДНК. Например, жидкая природа конденсатов RMM позволяет им специфическим образом задействовать или же исключать из хода процесса определенные факторы репарации ДНК.
Работа Claeys Bouuaert с соавт. знаменует начало удивительной «фазы» исследований фундаментальных механизмов мейотической рекомбинации. В сочетании с неуклонным прогрессом понимания строения и функционирования мейотических хромосом создается основа для будущих научных достижений, таких как реконструирование мейотических разрывов ДНК in vitro и рекомбинация гомологичных хромосом.
