
Разнообразие, развитие и регуляция кишечных нейронов

В 2021 году анализ транскриптов кишечника мыши и человека улучшил понимание клеточного состава, развития и микроокружения кишечной нервной системы (КНС). Влияние КНС на соотношения регуляторных T-лимфоцитов способствовало улучшению знаний о зависимости между КНС, иммунной системой и микробиотой.
▶ Еще по теме: Инфографика: Синдром раздраженного кишечника
Будучи уникальным среди внутренних органов, желудочно-кишечный тракт содержит собственную внутреннюю нервную систему — кишечную (КНС), — обеспечивающую автономный контроль перистальтики, кровотока и секреции [1]. Наличие более системной роли КНС можно предположить исходя из ее связи с иммунной системой, микробиотой, прочими внутренними органами и мозгом [1]. Тем не менее, из-за сложной анатомической структуры исследования функции КНС и ее развития отстают от таковых для центральной нервной системы (ЦНС). У человека КНС состоит из > 300 000 миллионов клеток, которые распределяются на протяжении > 5 м длины желудочно-кишечного тракта. Более того, в отличие от ЦНС, в которой анатомическое расположение связано с функцией клеток, КНС компартментализуется в малые неупорядоченные ганглии, в которых перемешаны различные типы клеток. Хотя функциональные кишечные нейроны были описаны так же, как и гетерогенная кишечная глия, неоднозначность молекулярных маркеров затруднила изучение полной сложной клеточной структуры.
В серии недавних анализов транскриптомов клеточный состав КНС начали определять и в ювенильных, и во взрослых стадиях [2–6]. В 2021 году классификация подтипов нейронов была усовершенствована [5] и использована для определения механизмов формирования идентичности нейронов в ходе развития КНС. Морарач с соавт. [5] определили 12 различных по молекулярным характеристикам классов нейронов мышечной оболочки кишечника, доказав их существование методами иммуногистохимии и описав как предполагаемые возбуждающие и тормозные мотонейроны, вставочные и чувствительные нейроны. Примечательно, что Nmu (кишечный иммунорегулятор [1]) рассматривается как истинный сенсорный класс, что предполагает наличие потенциального сопряжения между химической чувствительностью и способностью запускать иммунные реакции. Второй предположительно сенсорный класс не обладает классической морфологией клеток Догеля II типа (гладкие мультиполярные клеточные тела), но специфическая экспрессия Piezo2 указывает на их механочувствительность. Аморфная структура КНС отражает ее происхождение из стволовых клеток нервного гребня, которые проникают в кишечник на ранних стадиях развития и затем асинхронно мигрируют, пролиферируют и дифференцируются. Вопрос о том, как нейроны приобретают специфические характеристики в отсутствие пространственно определенных пулов стволовых клеток (сходных с прогениторными доменами в нервной трубке), остается нерешенным. С помощью построения карт профилей разных классов нейронов в развивающейся КНС (на 15,5 и 18,5 дни эмбрионального развития [E15.5 и E18.5]) в ходе исследования был открыт удивительный механизм развития. Только два класса нейронов формировались в ходе нейрогенеза (предположительно возбуждающие и тормозные моторные нейроны), в то время как прочие классы, по-видимому, возникают путем последующей дифферeцировки на постиитотических этапах (рис. 1a). Такой пошаговый процесс расхождения представляет собой концептуально новый способ возникновения разнообразия подтипов нейронов и требует дальнейшего исследования механизмов и молекулярных характеристик. С помощью оценки регуляторов транскрипции и передачи сигнала, определяемых в рамках транскриптома, в настоящее время будут специфичные для класса молекулярные программы; это откроет путь для создания эффективных протоколов дифференцировки для нужд регенеративной медицины на основе клеточных технологий или моделирования заболеваний. Например, Морарач с соавт. [5] наряду с авторами еще одного исследования [6] определили Pbx3 и Tbx3 как регуляторы развития тормозных мотонейронов — класса, число представителей которого уменьшено при ахалазии, диабете и болезни Чагаса.
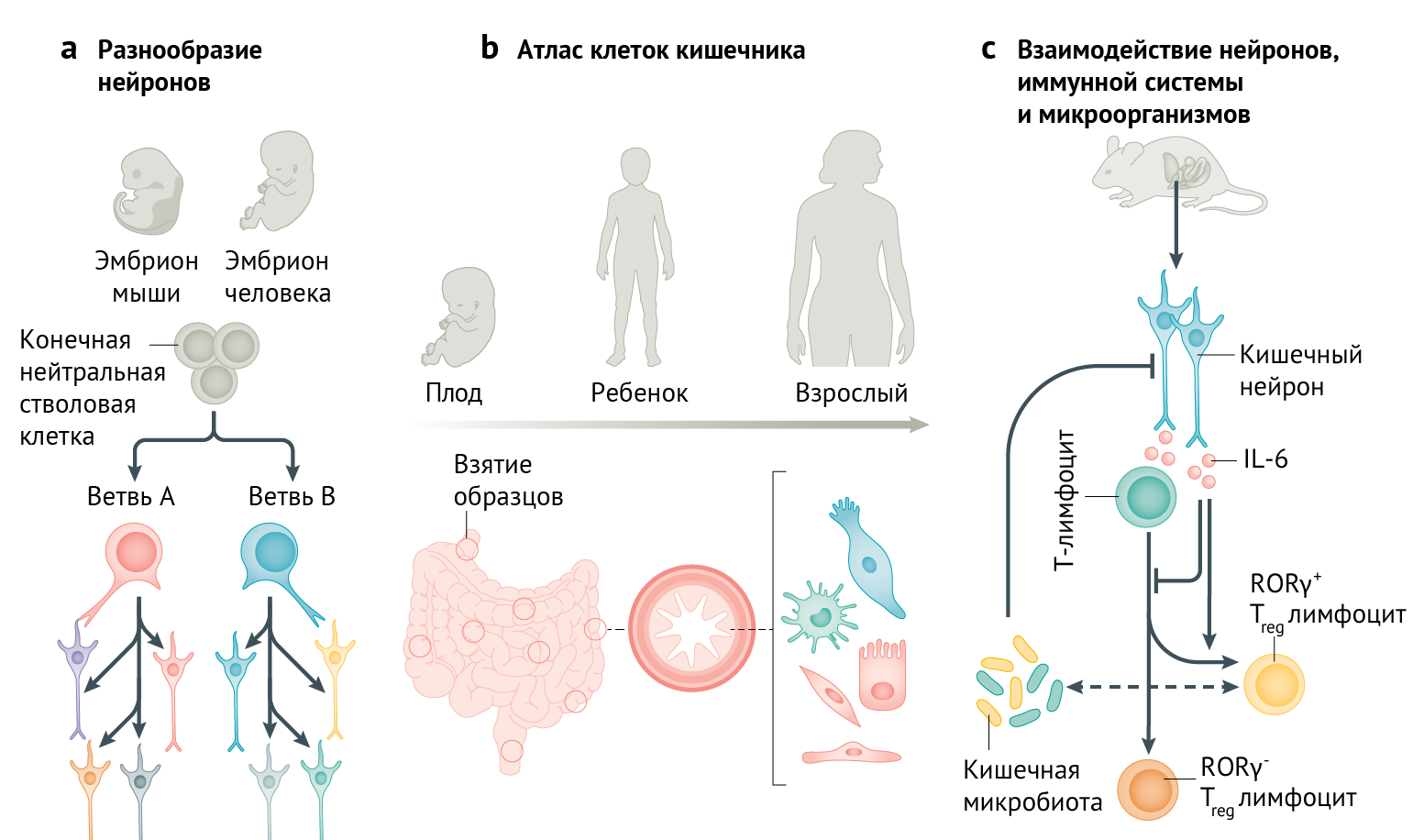
b | Всеобъемлющий атлас клеточного ландшафта на протяжении кишечного тракта человека включает разнообразное сочетание мезенхимных, эпителиальных, эндотелиальных, иммунных клеток и эритроцитов на стадии плода, ребенка и взрослого [7].
c | Реагирующие на микроорганизмы кишечные нейроны влияют на баланс количества и фенотипов регуляторных T-лимфоцитов (Treg) в согласованном трехчастном цикле регуляции кишечной толерантности [10].
Атласы КНС мышей формируют отличную базу для экспериментальных исследований, но не идеальны для достижения конечной цели — понимания точных ролей, которые КНС человека играет в здоровом организме и при патологиях. К счастью, за последние несколько лет было опубликовано несколько атласов транскриптомов единичных клеток кишечника плода, детей и взрослых (включая КНС) [3, 6–9]. На сегодняшний день наиболее всеобъемлющий атлас кишечника авторства Элмениате с соавт. [7] представляет монументальную характеристику 428 000 клеток во времени (6–17 недели после зачатия; детские и взрослые стадии) и пространстве (дo 11 регионов кишечника) (рис. 1b). Всего было охвачено 17 405 развивающихся клеток в период от раннего до среднего нейрогенеза (у мыши эквивалентный E12,5–E18,5). Обнаружено, что нейробласты дифференцируются на две основные нейронные ветви, характеризующиеся консервативными транскрипционными факторами [5] (A: ETV1 и B: BNC2), c последовательным появлением классов нейронов. Таким образом, формирование нейронного разнообразия в КНС человека, по-видимому, идет в соответствии с концептуальным принципом, выявленным у мышей (рис. 1a). Гены, повышающие риск болезни Гиршпрунга, демонстрируют дивергентную экспрессию — только RET в нейронах ветви A, а также ZEB2 и EDNRB с наибольшей экспрессией в толстой кишке — области, наиболее страдающей при болезни Гиршпрунга. Важно, что в атласе, помимо 133 описанных типов клеток и состояний, также уделено внимание клеточному контексту КНС, отличного от нейтрального происхождения. Например, важными источниками GDNF и EDN3 (лиганды сигнальных путей, изменяющихся при болезни Гиршпрунга) были гладкомышечные клетки и интерстициальные клетки Кахаля. Клетки вблизи КНС, включая различные эпителиальные, мезенхимные, эндотелиальные, иммунные и стромальные подтипы (рис. 1b). Атлас клеток кишечника человека [7] (доступен на gutcellatlas.org) наряду с другими выдающимися атласами [3, 6, 8, 9] представляет собой отличный источник для расшифровки клеточных и молекулярных коннектомов в физиологии желудочно-кишечного тракта.
КНС не может работать изолированно и сильно взаимосвязана с другими клеточными системами кишечника. В частности, появилась точка зрения, согласно которой нервная и иммунная системы кишечника эволюционировали совместно для согласованной работы по поддержанию клеточного гомеостазa и защиты [1]. В 2021 году Ян с соавт. [10] получили новые интересные данные о роли КНС в качестве ключевого регулятора состава иммунных клеток. Авторы обнаружили специфическую способность выделенных кишечных нейронов in vitro ингибировать дифференцировку регуляторных T-лимфоцитов (iTreg) путем обеспечения формирования подгруппы клеток RORγ+. В ходе системного изучения, включающего фракционирование нейрального супернатанта, определение сигнальных путей в Т-лимфоцитах, анализ взаимодействия рецепторов и лигандов и опосредованное антителами ингибирование, в качестве основного эффектора КНС был описан IL-6. Противоположный фенотип (увеличенный пул Treg; сниженный — подтипа RORγ+) был выявлен у мышей с нейрально-специфическим нокаутом Il6, что подтверждает этот механизм in vivo. Наконец, введение Clostridium ramosum, — бактерии-коменсала и известного индукторa RORγ+ Treg-клеток, в безмикробных мышей снижало плотность волокон КНС, экспрессию ряда нейромедиаторов, а также приводило к задержке экспрессии IL-6. Хотя иммунорегуляторная активность IL-6 известна давно, происхождение КНС неожиданно делает КНС центром внимания при исследовании дифференцировки иммунных клеток. Особенно сильно задействованы нейроны NOS1+ мышечной оболочки желудочно-кишечного тракта, но возможная роль нейронов подслизистой основы, которые располагаются ближе к Treg-клеткам и микроорганизмам, не изучалась, возможно, вследствие их недостаточной базовой характеристики. Действительно, обычным ограничением современных атласов КНС является то, что они фокусируются на типах клеток мышечной оболочки, и для выявления клеточного разнообразия внутри подслизистых сплетений необходимы дальнейшие исследования. Тем не менее, трехкомпонентное взаимодействие между кишечными нейронами, Treg-клетками и микроорганизмами кишечника (рис. 1c) позволяет предположить, что онтогенетическое, генетическое или средовое нарушение активности КНС и ее состава может изменять равновесие толерантности, взаимодействие между микроорганизмами и хозяином, а также течение воспалительных заболеваний. Таким образом, введение транскриптомики и нейроиммунологии в изучение КНС значительно расширило наши знания о строении кишечных нейронов, их разнообразии и взаимосвязи с другими клеточными системами. Проведение новых исследований на основе этого фундамента может предоставить новые возможные стратегии, такие как контроль или регенерация специфических типов кишечных нейронов, для предотвращения или лечения заболеваний желудочно-кишечного тракта.
