
Открыт новый уровень репрессии генов у человека

Белковый комплекс под названием риксосома помогает осуществлять деградацию транскриптов РНК, которые сохраняются после окончания экспрессии генов. Это открытие указывает на различные роли риксосомы в регуляции активности хроматина у разных видов.
После окончания экспрессии генов компоненты транскрипционного аппарата наряду с РНК-транскриптами могут оставаться поблизости от ДНК. Пути деградации транскриптов и репрессии генов в клетках человека известны, однако до недавнего времени эти пути не удавалось напрямую связать между собой. В журнале «Nature» Жоу с соавт. [1] описывают способ удаления РНК из неактивных человеческих генов, который запускается ключевыми участниками аппарата генной репрессии.
У многоклеточных организмов ДНК упакована в структуру под названием хроматин, который состоит из ДНК и гистоновых белков. В то время как ДНК несет генетическую информацию, белки-гистоны содержат информацию другого типа — гистоновый код [2] — различные химические модификации, которые передают специфическую для данного типа клеток информацию о том, какие гены должны быть активированы, а какие — репрессированы. При репрессии генов в определенном типе клеток они укладываются в компактную хроматиновую структуру, называемую факультативным гетерохроматином. В нем в изобилии присутствует — и считается его характерной чертой — набор репрессорных модификаций гистонов.
Среди белков, которые создают такие репрессорные модификации, комплексы PRC1 и PRC2. PRC подавляют синтез РНК в факультативном гетерохроматине с помощью нескольких механизмов: они маркируют гены для репрессии, запускают компактизацию хроматина, предотвращают связывание с хроматином факторов, активирующих генную экспрессию и, возможно, также предотвращают добавление ряда активирующих модификаций гистонов [3].
Но как уже синтезированная РНК удаляется из факультативного гетерохроматина, оставалось неясным. Это фундаментальный вопрос, поскольку ассоциированная с хроматином РНК связана со структурой хроматина [4, 5], стабильностью генома [6] и его функцией [7].
Жоу с соавт. решили изучить возможную роль белкового комплекса под названием риксосома в путях репрессии генов в клетках человека. Авторы исследовали две ферментативные функции риксосомы: первая — роль эндонуклеазы, которая расщепляет РНК, вторая — роль киназы, которая присоединяет фосфатные группы к расщепленной молекуле, помечая ее для деградации [8]. Ранее было показано, что эти активности риксосомы участвуют в создании репрессорных модификаций гистонов в клетках дрожжей [9].
Авторы показали, что риксосома колокализуется с комплексами PRC в факультативном гетерохроматине клеток человека и напрямую взаимодействует с PRC1. В связи с этим исследователи создали мутантную форму PRC1, неспособную связываться с риксосомой. В клетках, несущих мутантный белок, опосредованная PRC модификация гистонов происходила нормально, однако риксосома не могла локализоваться в факультативном гетерохроматине.
Далее Жоу с коллегами проанализировали клетки, в которых мутации риксосомы подавляли ее рибонуклеазную или киназную активность. В обоих случаях экспрессировался набор генов, который в норме репрессирован в факультативном гетерохроматине.
В совокупности полученные авторами данные говорят в пользу следующей модели. Сначала PRC1 рекрутирует риксосому в факультативный гетерохроматин. Далее риксосома используют свою эндонуклеазную и киназную активность для разрезания РНК и подготовки ее к деградации. Наконец, другой белок — экзорибонуклеаза XRN2 — заканчивает работу, расщепляя фосфорилированную РНК. Таким образом, PRC в клетках человека запускают каскад молекулярных событий, которые в конечном счете селективно устраняют РНК из факультативного гетерохроматина (рис. 1b).
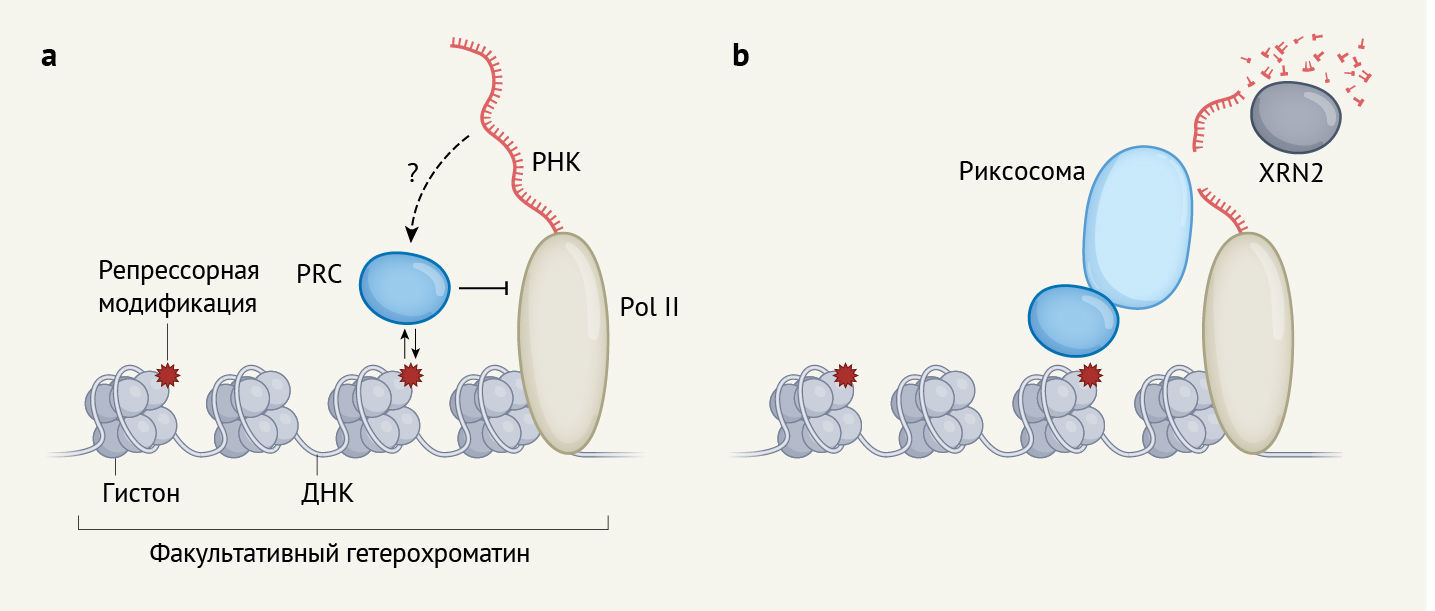
b | Жоу с соавт. показывают, что PRC также инициируют привлечение к факультативному гетерохроматину комплекса под названием риксосома. Риксосома расщепляет РНК, синтезированную Pol II, и подготавливает ее к деградации с участием другого фермента, XRN2.
У дрожжей ферментативная активность риксосомы необходима для правильной модификации гистонов в гетерохроматине [9]. Но они отличаются от тех, которые осуществляют PRC, a гетерохроматин дрожжей сильно отличается от факультативного гетерохроматина человека, поскольку последний эволюционировал для поддержания различных транскрипционных программ в разных типах клеток. Обширно изучаемые штаммы дрожжей фактически утратили PRC в ходе эволюции [3]. Таким образом, риксосомы дрожжей и человека оказались вовлечены в два различных пути регуляции хроматина.
Риксосома также участвует в образовании рибосомы — клеточного аппарата синтеза белка [8]. Однако Жоу с соавт. показали, что описанный ими путь не зависит от биогенеза рибосом. Tаким образом, в ходе эволюции назначение риксосомы неоднократно изменилось для выполнения различных функций в разных процессах, происходящих в ядре. Есть большой соблазн спекулировать, что риксосома также может принимать участие и в других путях, связанных с хроматином.
Множество PRC взаимодействует с тысячами транскриптов [10], и связанная РНК, по-видимому, регулирует активность PRC. Для объяснения того, как может происходить такая регуляция, были предложены разнообразные модели, но точные молекулярные механизмы остаются предметом изучения [11]. Работа Жоу с коллегами подразумевает, что опосредованная РНК регуляция PRC работает в обе стороны.
PRC регулируют друг друга с помощью сложных петель положительной обратной связи, в которой задействованы репрессорные модификации гистонов. Это делает трудным разделение прямых и косвенных эффектов при изучении регуляции репрессии генов под действием PRC. Эксперименты Жоу с соавт. указывают на субъединицу PRC1 в качестве ключевой детерминанты привлечения риксосомы к факультативному гетерохроматину. Менее изучено то, как риксосому регулирует PRC, хотя авторы определили физические взаимодействия между комплексами. Существует по крайней мере шесть подтипов комплекса PRC1 и два подтипа PRC2, у каждого из которых различаются субъединичный состав и функции [3]. Чтобы определить подтипы PRC1 (и, возможно, PRC2), которые рекрутируют риксосому к хроматину, потребуется еще больше работы. Структурные исследования могут прояснить, как при взаимодействии PRC и риксосомы эти массивные комплексы справляются со своими задачами в контексте хроматина.
Накапливающиеся данные указывают на РНК как на центральный остов, определяющий структуру [9] и компартментализацию хроматина [5]. Новообретенное знание о способности PRC запускать деградацию РНК вместе с их известной ролью в модификации гистонов и компактизации хроматина подтверждает значение этих комплексов как ключевых двигателей для организации трехмерной структуры и функционирования факультативного гетерохроматина.
