
Биология вариантов SARS-CoV-2: избегание иммунной системы, передача и приспособленность
Аннотация
В конце 2020, спустя почти год циркуляции в человеческой популяции, коронавирус острого респираторного синдрома 2 (SARS-CoV-2) претерпел крупное скачкообразное изменение в адаптации к человеку. Такие сильно мутировавшие формы SARS-CoV-2 имели повышенную способность к передаче по сравнению с предыдущими вариантами и были названы «вариантами, вызывающими обеспокоенность» (ВВО). Обозначенные как «альфа», «бета», «гамма», «дельта» и «омикрон», ВВО возникли независимо друг от друга, и каждый из них по очереди становился преобладающим — в отдельных регионах или по всему миру, — выигрывая конкуренцию с прочими вариантами. Успех каждого ВВО по сравнению с предыдущим доминирующим штаммом обеспечивался изменениями внутренних функциональных свойств вируса и различной степенью изменений антигенности вируса, придающих ему способность уклоняться от примированного иммунного ответа. Связанная с ВВО повышенная приспособленность вируса является результатом сложного взаимодействия биологии вируса и изменений иммунитета человека вследствие как вакцинации, так и ранее перенесенной инфекции. В настоящем обзоре обобщены литературные данные об относительной трансмиссивности и антигенности вариантов SARS-CoV-2, роли мутаций в сайте расщепления Spike-белка фурином и мутаций прочих белков, потенциальной важности рекомбинации для успеха вируса и эволюции SARS-CoV-2 в контексте Т-клеток, врожденного и популяционного иммунитета. SARS-CoV-2 демонстрирует сложную взаимосвязь между антигенностью, передачей и вирулентностью, что имеет непредсказуемые последствия для будущей траектории и бремени болезни COVID-19.
Введение
С момента своего первого появления в городе Ухань в декабре 2019 года коронавирус тяжелого острого респираторного синдрома 2 (SARS-CoV-2) стал причиной более 641 миллиона случаев заболевания COVID-19 и более 6,6 миллиона смертей по состоянию на декабрь 2022 года [1]. SARS-CoV-2 (наряду с SARS-CoV, причиной SARS) является представителем вида коронавирусов, связанных с тяжелым острым респираторным синдромом, единственным в подроде сарбековирусов, в основном обнаруживаемых у подковоносых летучих мышей [2]. Как и другие коронавирусы, SARS-CoV-2 обладает большим РНК-геномом, состоящим примерно из 30 000 нуклеотидов, репликация которых опосредована РНК-зависимой РНК-полимеразой (RdRP) и связанным корректирующим ферментом экзорибонуклеазой (ExoN). В сочетании с прерывистым характером транскрипции коронавируса это привело к появлению коронавирусов с высокой скоростью рекомбинации, вставок и делеций, а также точечных мутаций (хотя скорость ниже, чем для других РНК-вирусов из-за корректировки при копировании матрицы), как было описано ранее [3]. Успешность образующихся новых генетических вариантов, хотя и подвержена стохастическим процессам, будет сильно зависеть от естественного отбора — в частности, положительного отбора, связанного с мутациями, которые полезны для несущего их вируса.
SARS-CoV-2 оказался сильным патогеном для человека, а также универсалом с точки зрения тропизма к хозяину, вызывая инфекции у различных видов млекопитающих; в качестве примеров можно привести инфекции у норок на зверофермах [4], стабильный резервуар в белохвостых оленях [5, 6] и случайные инфекции многих других видов животных [7]. После появления SARS-CoV-2 в человеческой популяции его эволюция в течение первых месяцев характеризовалась ограниченной адаптацией и фенотипическими изменениями по сравнению с теми, что происходили в ходе дальнейшей эволюции [8]. Первое заметное изменение — замена одной аминокислоты в Spike-белке (D614G) — возникло в начале пандемии и обеспечило преимущество в росте примерно на 20% по сравнению с предыдущими вариантами [9]. Линия, определяемая D614G (линия PANGO10 B.1), быстро стала доминирующей в Европе, что стало ранним указанием на усиление трансмиссивности SARS-CoV-2 среди людей. Как авторы обзора описывали ранее [3, 11], с октября 2020 года начали появляться новые, сильнее мутировавшие варианты SARS-CoV-2. Они отличались большим количеством несинонимичных мутаций, главным образом, в Spike-белке — особенно в случае «омикрона» — и различными фенотипическими свойствами, включая измененную трансмиссивность и антигенность. На сегодняшний день Всемирная организация здравоохранения и национальные агентства общественного здравоохранения объявили вариантами, вызывающими обеспокоенность (ВВО), пять штаммов SARS-CoV-2 на основании их существенно измененной трансмиссивности или способности избегать иммунитета, что требует тщательного мониторинга. Каждый ВВО демонстрировал преимущества передачи по сравнению с предыдущими вариантами и стал доминирующим либо регионально — в случаях штаммов «альфа» (линия PANGO10 B.1.1.7), «бета» (B.1.351) и «гамма» (P.1) — в Европе, Южной Африке и Южной Америке, соответственно, — или глобально, в случаях «дельты» (подлинии B.1.617.2/AY) и многих подлиний «омикрона» (подлинии B.1.1.529/BA, такие как BA.1, BA.2 и BA.5).
Вставка 1. Происхождение вариантов SARS-CoV-2, вызывающих обеспокоенность, и комплекс «омикрон»
Варианты коронавируса острого респираторного синдрома 2 (SARS-CoV-2), вызывающие обеспокоенность (ВВО), несут ряд отличительных свойств. Одно из наиболее интригующих — относительно большое филогенетическое расстояние, часто с отсутствием генетических промежуточных звеньев до их обнаружения [67, 193, 194]. Вариант «альфа» был впервые обнаружен в Великобритании в период низкого уровня циркуляции вируса и беспрецедентного эпиднадзора за SARS-CoV-2 [67, 131]. Вариант «дельта» впервые выявлен в Индии в апреле 2021 года [195], а затем был связана с новой волной эпидемии в Великобритании [196] и образовал несколько подлиний, некоторые из которых были даже более трансмиссивными, чем родительский штамм «дельта», — например, AY.4.2 [197]. Сходным образом, комплекс вариантов «омикрон» имеет значительную длину филогенетических ветвей и даже больше мутаций, чем предшествующие ВВО [43, 44].
Отсутствие промежуточных последовательностей привело к созданию нескольких гипотез происхождения ВВО [198]: (1) циркуляция в регионах с грубым шагом географической дискретизации; (2) скрытая циркуляция в животном резервуаре после обратного зооноза раннего варианта; (3) эволюция в рамках хронической инфекции у хозяина с подавленным иммунитетом или у нескольких хозяев. Хотя существуют данные о появлении видов животных, служащих стабильными резервуарами для SARS-CoV-2, — особенно у разводимых на фермах норок и белохвостых оленей [4, 5, 186] — в настоящее время из этих сценариев появления новых вариантов «гипотеза хронической инфекции» подтверждается лучше всего [67, 183]. Известно, что хронические инфекции способствуют возникновению мутационных профилей, очень похожих на профили ВВО [113, 184, 199–201]. Есть также данные об ограничениях дальнейшей передачи хронических инфекций, вызывающих локальные вспышки [113, 182]. Антигенное расстояние ВВО, особенно для «омикрона», может объяснить длительная эволюция в присутствии постоянного давления субнейтрализующих антител.
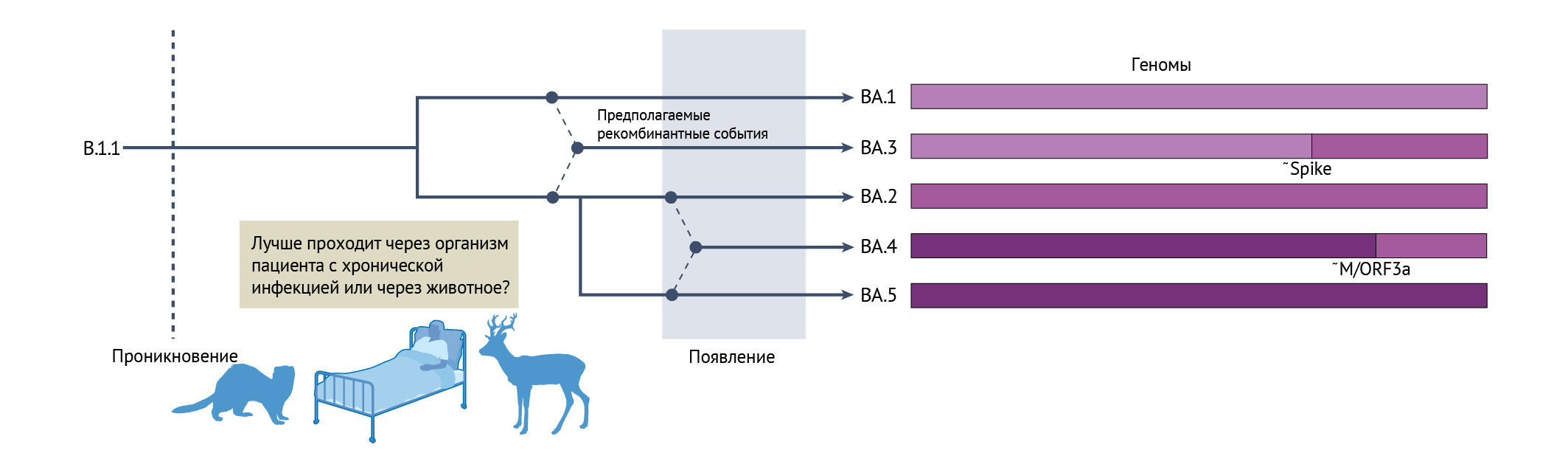
В отличие от других ВВО, у «омикрона» в ходе эволюции возникло удивительное разнообразие до того, как он был впервые обнаружен. В настоящее время выделяют пять основных линий (BA.1, BA.2, BA.3, BA.4 и BA.5 [43] (см. рисунок) с еще большим числом обнаруженных подлиний, которые накапливают дальнейшие изменения антигенных свойств [157]. BA.1 вызвала глобальную инфекционную волну в конце 2021 года, но к началу 2022 года его сменила BA.2. В апреле 2022 года были выявлены дополнительные подлинии BA, BA.4 и BA.5, а по состоянию на сентябрь 2022 года BA.5 встала во главу новой общемировой волны «омикрона» [43]. Линии «омикрона» связаны друг с другом сложным родством, которое, вероятно, связано с несколькими событиями рекомбинации внутри ВВО до обнаружения [43, 44]. Например, BA.4 и BA.5 содержат почти идентичные 5'-концы генома до гена M, но после этого демонстрируют большое расхождение, что указывает на недавнее событие рекомбинации. Сходным образом, BA.3 предположительно может быть рекомбинантным потомком предковых вирусов BA.1 и BA.2 [44]. На рисунке показан гипотетический филогенетический источник комплекса «омикрон», демонстрирующий один возможный паттерн рекомбинации среди подлиний BA.1, BA.2, BA.3, BA.4 и BA.5. Названия белков указывают приблизительные (~) потенциальные точки разрыва.
Вопреки ожиданиям, что после появления в человеческой популяции вирусы быстро адаптируются к хозяину [12, 13], анализ отбора показывает, что в начале пандемии SARS-CoV-2 не обладал заметными уровнями адаптации [14]. Впоследствии стало ясно, что SARS-CoV-2 является универсальным вирусом, способным использовать различные мембранные белки ангиотензинпревращающего фермента 2 (AПФ2) млекопитающих для проникновения в клетку [15], что позволяет инфицировать широкий круг млекопитающих [14, 16]. Сарбековирусы часто передаются между различными видами подковоносых летучих мышей [17], а также другими видами животных, у которых он способен связываться с АПФ2 (предполагаемый предковый признак сарбековирусов [18]), включая и людей. Spike-белок SARS-CoV-2 обладает важными свойствами, которые необходимы для эффективной передачи от человека к человеку, — в частности, способностью к связыванию с человеческим АПФ2 и многоосновным сайтом расщепления фурина (FCS) на стыке S1–S2 [19, 20]. В настоящее время FCS SARS-CoV-2 S1-S2 уникальна среди сарбековирусов, хотя аналогичные последовательности встречаются и у других бета-коронавирусов.
Для проникновения SARS-CoV-2 в клетки дыхательных путей требуется опосредуемое фурином расщепление FCS, которое делает возможным слияние мембран. Таким образом, FCS служит ключевым фактором, определяющим высокую скорость передачи SARS-CoV-2, способствуя его эффективному распространению среди людей [19, 21]. Дальнейшая оптимизация FCS дикого типа в ходе пандемии привела к усиленному расщеплению фурином Spike-белков у вирусов «альфа» и «дельта» [22–25]. Считается, что в сочетании с другими мутациями, особенно теми, которые усиливают связывание АПФ2 [26, 27], мутации, оптимизирующие расщепление фурина, способствовали повышенной трансмиссивности ВВО «альфа» и «дельта», которая оказалась повышена на 65% и 55%, соответственно, относительно тех вариантов, которые они вытеснили [28–30]. В отличие от «альфы» и «дельты», эволюционный успех варианта «омикрона» не связан с оптимизацией расщепления фурина. Скорее, «омикрон» характеризуется измененным фенотипом проникновения [31, 32] в совокупности со значительной способностью избегать иммунитета [31–34], что позволяет ему эффективно заражать вакцинированных или ранее инфицированных людей. Хотя трансмиссивность в наивной популяции в значительной степени определяется внутренними свойствами вируса, все более сложный иммунный ландшафт, в котором сейчас циркулирует SARS-CoV-2, означает, что основной движущей силой успешности штамма становится ускользание от антител (в противоположность усилению трансмиссивности только за счет биологии самого вируса — черте, которую сложно оптимизировать сильнее, нежели это удалось «омикрону»). До появления «омикрона» (вставка 1) каждый из преобладающих вариантов эволюционировал из собственных предшественников, но не друг из друга. Напротив, сейчас последовательные волны вызываются подлиниями «омикрона» (например, BA.5 — одна из подлиний BQ.1 и BA.2.75 — подлиния BA.2). Стоит отметить, что возможно появление еще не известного варианта, — возможно, рекомбинантного (вставка 2) — с высокой трансмиссивностью, связанной со внутренней биологией вируса и новыми антигенными свойствами.
Вставка 2. Рекомбинанты SARS-CoV-2
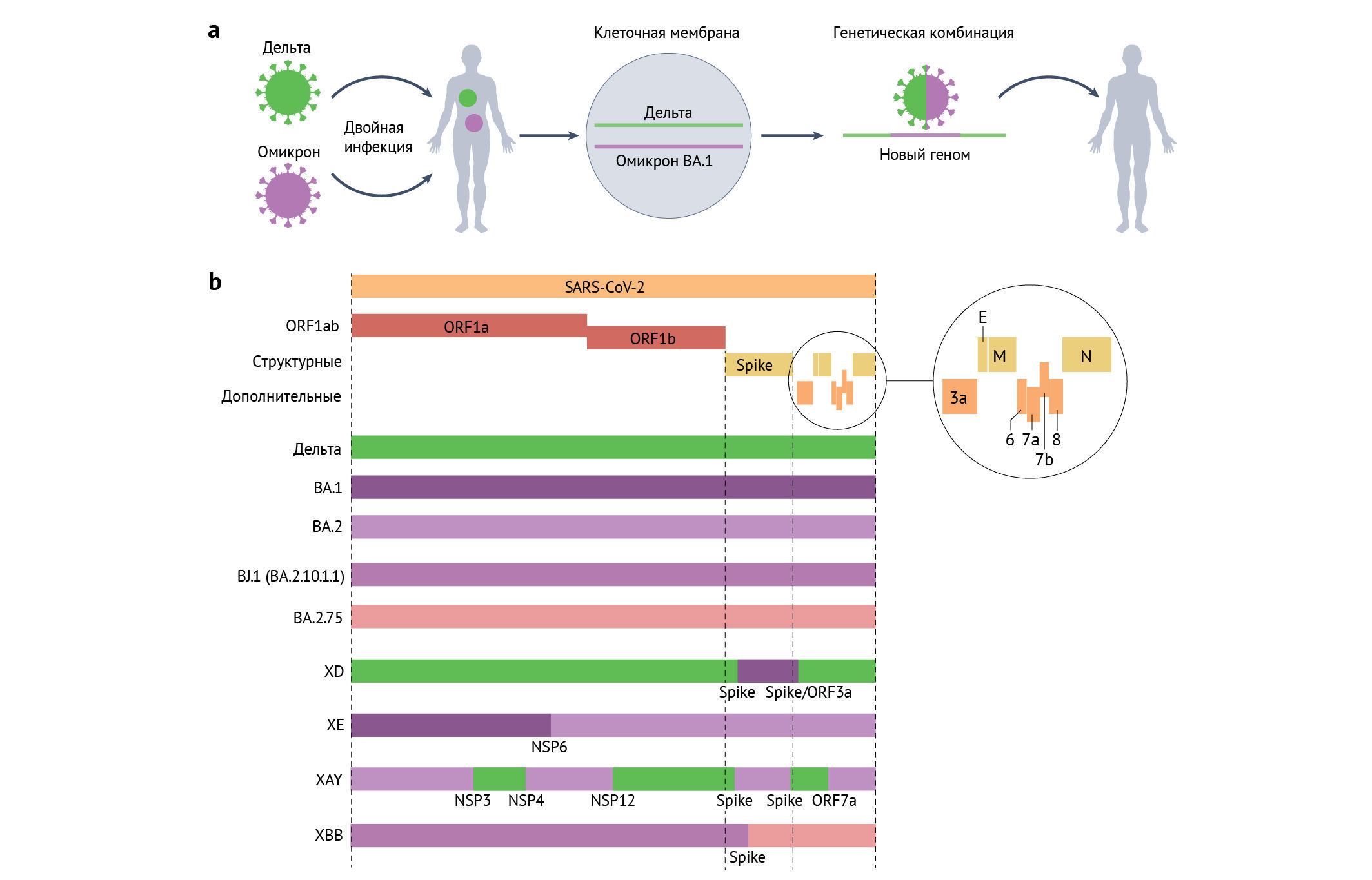
Когда два РНК-вируса совместно заражают одну и ту же клетку внутри организма, во время репликации генома существует высокая вероятность того, что полимераза переключится с одной матричной геномной последовательности на гетерологичный геном. В результате возникнет рекомбинантный вирус с частью генома от одного «родителя» и оставшейся геномной последовательностью от другого (см. рисунок, часть а). Несколько рекомбинантных линий были выявлены в разных местах и обозначены системой классификации PANGO (введена с 2020 года) [10] с добавлением префикса линии X-. Рекомбинации однозначно определяются в случае, когда генетически различные варианты, такие как два ВВО, в определенный период времени циркулируют совместно, что приводит к коинфекции. Например, первая линия, считающаяся рекомбинантной, — XA — была рекомбинантом между вариантом «альфа» (B.1.1.7) и линией, ранее циркулировавшей в Великобритании, B.1.177 [202]. Линия XC была обнаруженным в Японии рекомбинантом между «альфой» и «дельтой» [203].
С начала 2022 года количество выявленных рекомбинантов быстро увеличивалось, что, вероятно, было связано с высоким уровнем совместной циркуляции вариантов «дельта» и BA.1 или BA.1 и BA.2 во многих странах во время поэтапного снятия карантинных ограничений. Другим объяснением служит большая уверенность в определении рекомбинантов из-за более высокого расхождения последовательностей среди геномов «дельты», BA.1 и BA.2. Одним из примеров рекомбинанта является XD — рекомбинант «дельта» × BA.1, впервые обнаруженный в январе 2022 года во Франции [204]. XD имеет две точки разрыва генома, с остовом и частью N-концевого домена Spike-белка «дельты» и оставшейся частью — от BA.1 (см. рисунок, часть b). Было показано, что у трансгенных мышей, экспрессирующих человеческий ангиотензинпревращающий фермент 2 (AПФ2) под контролем промотора кератина 18, функционально XD имеет промежуточный фенотип патогенности между BA.1 и «дельтой», вызывая умеренную потерю веса; это позволяет предположить, что часть дифференциального фенотипа патогенности для грызунов у вариантов «дельта» и BA.1 картируется вне Spike-белка.
Вторым примечательным рекомбинантом в Великобритании является XE BA.1 × BA.2, который впервые был обнаружен в Англии 19 января 2022 года. До того, как его сменил BA.5, XE включал более 2500 геномов, в основном из Великобритании. Предварительные данные предполагают умеренное возрастание темпов роста по сравнению с BA.2 [205]. XE содержит Spike и структурные белки от BA.2 и часть ORF1AB от BA.1.
Примечательна недавняя линия XAY — рекомбинант BA.2 × «дельта», который был впервые обнаружен в Южной Африке в мае 2022 года вместе с меньшим сестринским рекомбинантом, XBA [206]. Эти рекомбинанты имеют гораздо большее число точек разрыва, чем любой ранее известный кластер, а также множество уникальных мутаций, не обнаруженных в родительских линиях. Одним из возможных объяснений появления этих рекомбинантов служит хроническая инфекция вариантами «дельта» BA.2 и неоднократная рекомбинация (наряду с постепенным накоплением уникальных мутаций) перед повторным появлением в общей популяции. XBB — недавно возникший и быстро размножающийся рекомбинант между двумя линиями BA.2 второго поколения — BJ.1 (также известной как BA.2.10.1.1) и BA.2.75. Он содержит большое количество мутаций антигенного рецептор-связывающего домена; показано, что он плохо нейтрализуется сыворотками с антителами, которые ранее эффективно действовали на «омикрон».
Как уже упоминалось, специфические для варианта свойства могут картироваться как внутри, так и вне Spike-белка. Рекомбинанты, такие как XD, демонстрируют возможность возникновения вирусов с фенотипическими свойствами обоих родительских вирусов [204]. До быстрого появления размножающегося XBB рекомбинанты не оказывали большого влияния на течение пандемии, обычно возникая примерно в то время, когда новые варианты заменяли ранее доминирующие линии. Следует отметить, что такая рекомбинация — обычное явление среди сарбековирусов, заражающих подковообразных летучих мышей, о чем свидетельствуют обнаруженные высокие уровни рекомбинантных геномов и тенденция к замене Spike-белка. Это указывает на частые события антигенного сдвига [208]. Процесс рекомбинации, происходящий у человека с инфекцией, вызванной вариантами «омикрон» и «дельта» (см. рисунок, часть а). Карты геномов некоторых известных современных рекомбинантов (XD, XE, XAY и XBB) (см. рисунок, часть b)
E — белок оболочки; M — мембранный белок; N — белок нуклеокапсида; НСБ — неструктурные белки; SARS-CoV-2 — коронавирус острого респираторного синдрома 2.
Вне зависимости от того, возникнет ли полностью новый штамм, или же будущие вирусы разовьются из подлиний «омикрона» с новыми антигенными изменениями (что особенно вероятно, поскольку предыдущие ВВО больше не циркулируют в популяции), очевидно, что будут продолжать появляться новые штаммы SARS-CoV-2, обладающие уникальными комбинациями мутаций, и те из них, которые окажутся наиболее приспособлены, будут преобладать над прежними вариантами. На сегодняшний день успешные штаммы также демонстрируют вариации клинически значимых признаков, включая тяжесть заболевания, уклонение от иммунитета и чувствительность к терапевтическим средствам (в частности, к моноклональным антителам). Поэтому для общественного здравоохранения и клинического значения важно понять движущие силы приспособленности у SARS-CoV-2. Приспособленность варианта, то есть репродуктивный успех вируса, зависит от разнообразных факторов, которые определяют его способность заражать, размножаться внутри хозяев и распространяться между ними. В настоящем обзоре авторы рассматривают изученные мутации, для которых описано влияние на способность SARS-CoV-2 к заражению и передаче, и обсуждают способность вируса избегать Т-клеточный, врожденный или гуморальный иммунитет [11]. Spike-опосредованный гуморальный иммунитет рассмотрен в предыдущем обзоре [11].
Варианты SARS-CoV-2 и антигенное избегание
Рано возникшая проблема, касающаяся эволюции SARS-CoV-2, состояла в возможности возникновения вариантов с наличными антигенными свойствами, способных избегать иммунитета, приобретенного вследствие вакцинации или перенесенной инфекции; в качестве примера можно привести замену N439K в Spike-белке [35]. До усовершенствования в конце 2022 года в основе всех широко применяемых вакцин против COVID-19 лежал антиген Spike-белка ранних штаммов; в большинстве использовалась референсная последовательность Wuhan-Hu-1, образец которой был получен от инфекции линии B на Уханьском рынке морепродуктов — часто с мутациями, которые стабилизуют Spike-белок в конформации перед слиянием [36]. Хотя для «альфа»-штамма сообщалось об ограниченных антигенных изменениях [25, 37, 38], для вариантов «бета», «гамма» и «дельта» в лабораторных экспериментах было показано умеренное избегание индуцированных вакцинами антител и сыворотки выздоровевших пациентов [23, 37–39]. Тем не менее, эпидемиологические исследования показали, что эффективность вакцины против вариантов «бета» и «дельта» в значительной степени сохраняется [40–42]. Таким образом, несмотря на дизайн, в основе которого лежала очень ранняя последовательность Spike-белка, вакцины первого поколения против SARS-CoV-2 обеспечили замечательную защиту от тяжелых заболеваний и позволили большей части мира вернуться к подобию нормальности.
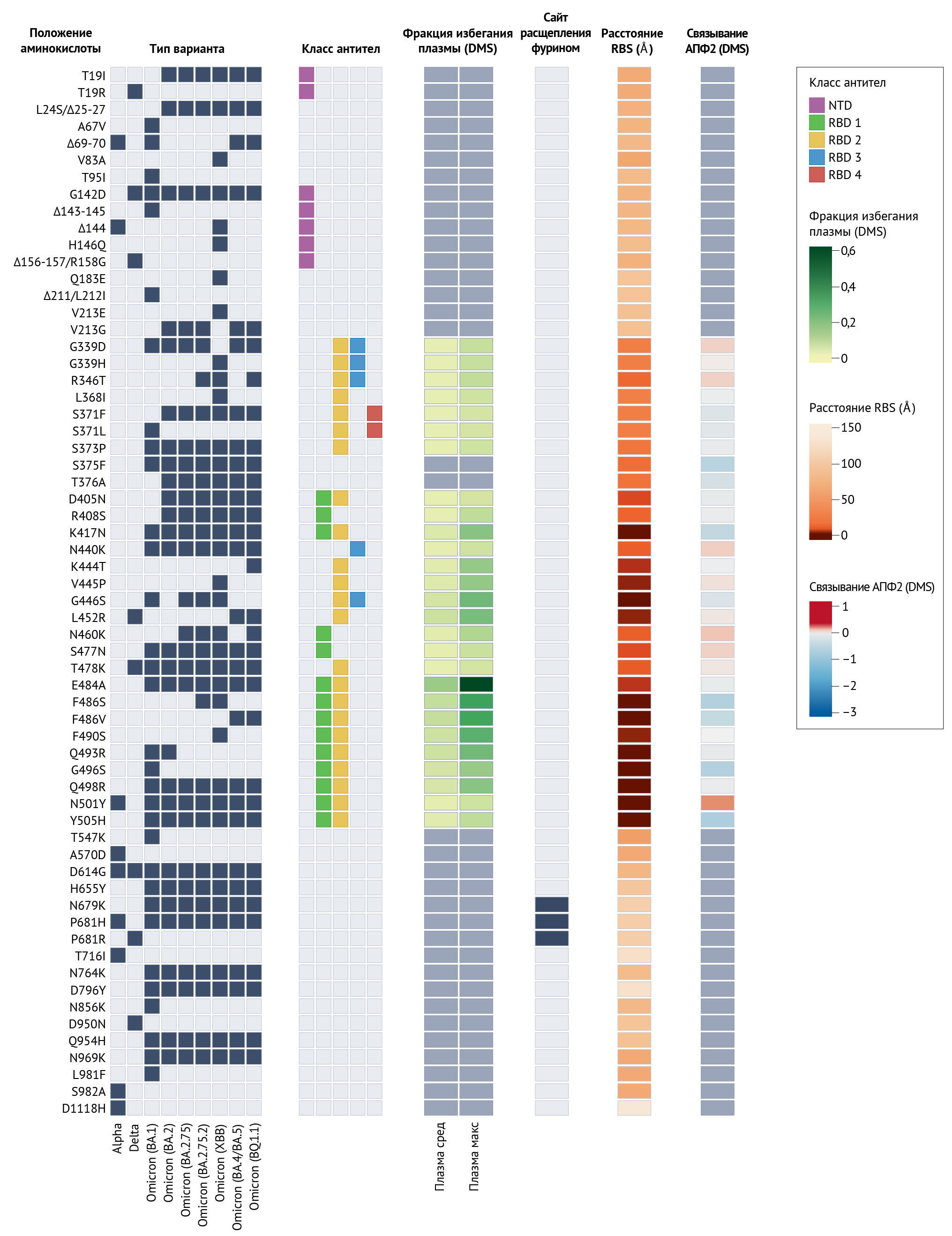
Комплекс «омикрон», состоящий из отдельных подлиний BA.1, BA.2, BA.3, BA.4 и BA.5 (вставка 1), способен заражать вакцинированных и ранее инфицированных людей, и выносит на первый план обсуждения стратегии борьбы с SARS-CoV-2 проблемы выбора последовательности для разработки вакцин и вопрос создания универсальных вакцин. Имея более 15 мутаций рецептор-связывающего домена (RBD) Spike-белка и ряд антигенных делеций и замен в N-концевом домене (NTD) [43, 44], BA.1, BA.2, BA.4 и BA.5 очень плохо нейтрализуются вакцинами первого поколения и антителами, полученными до появления «омикрона» (рис. 1). Кроме того, показано ускользание от подавляющего большинства современных терапевтических моноклональных антител; в настоящее время известно, что только бебтеловимаб — моноклональное антитело, направленное на RBD Spike-белка, — сохраняет свою эффективность против всех вариантов SARS-CoV-2 [31, 33, 43, 38, 45–50]. Этот крупный антигенный «сдвиг» привел некоторых исследователей к предположению, что линии «омикрона» следует рассматривать как отдельный штамм или серотип по сравнению с линиями до его появления [51, 52]. Важно отметить, что масштабы этого антигенного изменения отражены в данных о реальной эффективности вакцин против инфекций и симптоматических заболеваний [41, 53–57]. Для поддержания любой эффективности вакцины против «омикрона», которая снижается по мере снижения титров антител, необходимы бустерные дозы вакцин [41, 55]. Действительно, эффективность вакцины против тяжелой формы инфекции «омикроном» оставалась высокой через четыре месяца после бустерной дозы, а затем быстро снижалась, хотя снижение шло медленнее, чем после первичной вакцинации [58]. Из-за небольшой продолжительности защитного иммунитета против инфицирования «омикроном» при применении современных вакцин многие производители вакцин и ученые сосредотачиваются на вакцинах второго поколения, таких как моновалентные или бивалентные бустеры, специфичные для «омикрона» [59] (которые внедряются в настоящее время), вакцины с назальной доставкой для лучшей стимуляции иммунитета слизистых оболочек [60] или универсальные вакцины [61]. Как и в случае с сезонными коронавирусами человека, степень, в которой долгосрочный приобретенный иммунитет может предотвратить повторное заражение SARS-CoV-2, ограничивается сочетанием снижения уровня антител и антигенного дрейфа вируса — постепенного приобретения мутаций, позволяющих уклоняться от иммунитета [62, 63]. В дополнение к скачкообразным изменениям антигенности, эволюция вируса во время персистирующих инфекций позволила SARS-CoV-2 накопить множество мутаций в контексте одной или нескольких длительных инфекций, способствуя событиям антигенного сдвига, когда эти варианты продолжают инфицировать другие организмы (вставка 1).
FCS и появление вариантов SARS-CoV-2
В отличие от других известных сарбековирусов, SARS-CoV-2 обладает уникальной чертой — FCS в составе Spike-белка, который может расщепляться фурином. FCS, тем не менее, присутствуют у многих других бета-коронавирусов из подродов Embecovirus и Merbecovirus, таких как коронавирус человека OC43, коронавирус человека HKU1 и коронавирус средневосточного респираторного синдрома (рис. 2a). Показано, что у SARS-CoV-2 FCS необходим для оптимальной репликации вируса в клетках дыхательных путей человека [64], трансмиссивности [19, 21] и патогенности [65]. Фурин — протеаза хозяина — содержится преимущество в аппарате Гольджи, обеспечивая расщепление в ходе переноса вируса на клеточную поверхность [66]. Однако теперь понятно, что FCS ранних вариантов SARS-CoV-2 была субоптимальной и расщеплялась фурином неэффективно [19, 24, 65]. Интересно, что одна из ролей, описанных для ранней замещающей мутации в Spike-белке D614G, — умеренное усиление расщепления Spike-белка (подробно рассмотрено ранее [3]). Ряд последующих вариантов SARS-CoV-2 содержит мутации, прилегающие к FCS, которые увеличивают количество основных аминокислотных остатков — известного сайта узнавания фурина; например, «альфа», «мю» и «омикрон» содержат мутацию FCS P681H [44, 67, 68], которая, как предполагается, увеличивает активность расщепления фурином [19, 24, 65]. Более того, другая мутация в том же положении, P681R, усиливает репликацию и патогенность ВВО «дельта» [23, 30, 69] (рис. 2b). Следует отметить, что «омикрон» содержит P681H, а также дополнительную мутацию N679K44, которые вместе приводят к оптимизации FCS [19, 23–25, 32, 70]. Важно, однако, что оптимизация сайта расщепления фурином сама по себе не повышает трансмиссивность или репликацию SARS-CoV-2 и может мешать эффективной передаче вируса [27], указывая на то, что дополнительные мутации, наблюдаемые в этих вариантах, необходимы для оптимизации репликации и трансмиссивности.
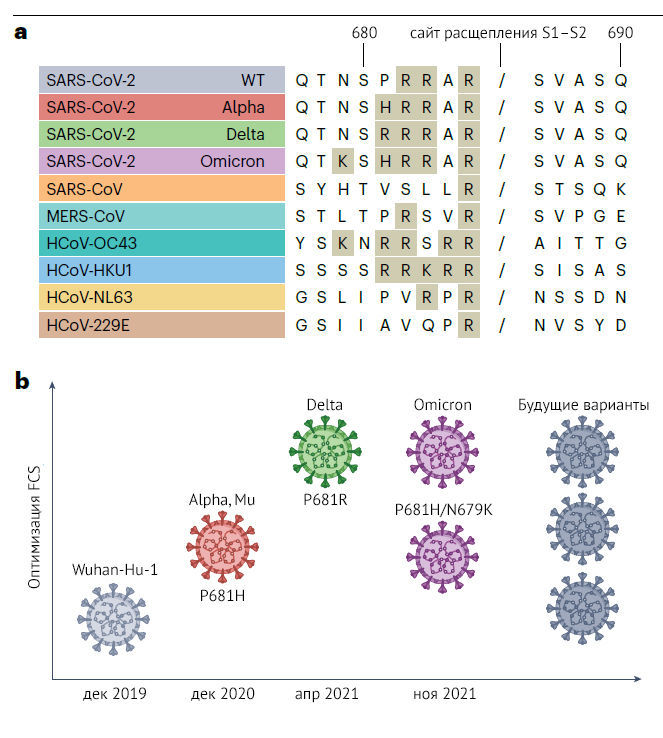
b. схематическое изображение предполагаемой относительной оптимизации сайта расщепления фурина S1–S2 (FCS) для рассматриваемых вариантов. Указаны мутации, влияющие на функцию FCS. Обратите внимание на то, что для «омикрона» получены несогласованные результаты. Уровень оптимизации FCS для будущих вариантов неясен. Данные о ВВО «альфа» из [19, 24, 25], данные о ВВО «дельта» из [22, 23, 69] и данные по ВВО «омикрон» из [32, 75].
Точный механизм, с помощью которого наблюдаемые мутации FCS усиливают его расщепление фурином, остается предметом дискуссий. Хотя есть достаточно убедительные доказательства того, что P681R напрямую усиливает взаимодействие с фурином и расщепление сайта S1–S2 [19, 24, 69], функциональные последствия P681H [71] менее ясны. С точки зрения механизма, вероятно, важно, что положение T678 Spike-белка рядом с остатком P681 может быть посттрансляционно модифицировано путем O-гликозилирования [72–74]. Известно, что О-гликозилированию способствуют нижележащие пролины; таким образом, альтернативным объяснением может быть то, что к потере гликозилирования, потенциально блокирующего сайт расщепления фурином и связанному с этим усилению расщепления приводит удаление P681, а не само по себе добавление гистидина.
Недавние исследования биологии «омикрона» поставили под сомнение гипотезу о том, что для повышения эффективности передачи вируса необходимо усиление расщепления Spike-белка. Все линии «омикрона» содержат P681H и N679K [43, 44], которые по отдельности или совместно усиливают расщепление сайта S1–S2 в Spike-белке дикого типа [32, 70]. Тем не менее, в контексте Spike-белка свидетельства такого улучшенного расщепления менее очевидны: в некоторых исследованиях обнаружено, что связь S1–S2 расщепляется хуже, чем в предыдущих ВВО [50, 75], в то время как в других показано, что эффективность расщепления сравнима с таковой у варианта «дельта» [32, 76, 77]. Многие группы описали, как «омикрон» может эффективно использовать альтернативный путь входа в клетку независимо от фенотипа расщепления.
Действительно, в то время как предыдущие ВВО, такие как «дельта», в значительной степени полагаются на праймирование слияния трансмембранной сериновой протеазой 2 (TMPRSS2) на клеточной поверхности, «омикрон», подобно SARS-CoV, также может эффективно праймироваться эндосомными протеазами, такими как катепсины [31, 32, 50, 75, 76, 78, 79]. Предполагается, что этот альтернативный механизм частично отвечает за снижение тяжести инфекции в случае «омикрона» — по крайней мере, на моделях грызунов [32, 75, 78, 80, 81] — из-за более низкой фузогенности и потенциально измененного тканевого тропизма со смещением инфекции от нижних дыхательных путей в сторону верхних. В нескольких исследованиях были предложены молекулярные механизмы для такой пониженной фузогенности и изменения пути проникновения, включая мутации в RBD Spike-белка «омикрона» [31, 82], H655Y [83] или мутации в домене S2 [31, 32, 76] — в частности, N969K32, хотя вопрос, сохраняется ли этот признак во всех линиях «омикрона», спорный. Тем не менее, линии «омикрона» продолжают демонстрировать высокую трансмиссивность, по крайней мере эквивалентную таковой у варианта «дельта» [85, 86], что подразумевает потенциальное разделение между эффективностью расщепления фурином и фузогенностью и их вкладом в эффективность передачи вируса.
Помимо сайта S1–S2, бета-коронавирусам также требуется расщепление протеазой второго сайта, известного как сайт S2'. После расщепления сайта S1–S2 и связывания с соответствующим рецептором сайт S2' экспонируется в домене S2 Spike-белка [87]. Расщепление S2' напрямую высвобождает пептид, необходимый для слияния мембран вируса и хозяина, которое и следует за этим процессом [88–90]. При проникновении SARS-CoV-2 в клетки дыхательных путей этот сайт расщепляется преимущественно сериновыми протеазами хозяина, такими как TMPRSS2, но есть и альтернативный путь, при котором расщепление осуществляют эндолизосомные катепсины [19, 91]. В ряде недавних статей высказывалось предположение, что вариации NTD SARS-CoV-2, особенно за счет ремоделирования внешних петель путем приобретения делеций или вставок, могут аллостерически влиять как на расщепление S1–S2, так и на расщепление S2', и, следовательно, на слияние [92–94].
В целом взаимосвязь между действием протеаз, эффективностью расщепления S1–S2, тропностью, патогенностью и трансмиссивностью SARS-CoV-2 очень сложная, и результаты исследований порой противоречивы. Таким образом, чтобы преодолеть эти пробелы в знаниях и полностью понять эту систему, необходима дальнейшая работа.
Прочие структурные и неструктурные белки и инфицирующая способность
В нескольких недавних исследованиях изучали последствия мутаций в структурных белках, помимо Spike, включая мембранные (М), белки оболочки (Е) и белки нуклеокапсида (N) белки. Линия B.1.1 определяется парой замен в белке N — R203K и G204R. Варианты, происходящие от B.1.1 (такие как «альфа», «гамма» и «омикрон»), унаследовали одни и те же мутации, тогда как у вариантов «бета» и «дельта» независимо возникли мутации с конвергентными функциональными свойствами R203M и T205I, соответственно. В частности, показано, что эти мутации повышают инфекционную способность вируса [95–97], хотя точный механизм действия остается спорным. C одной стороны, данные анализа по гену-репортеру вирусоподобных частиц [95] предполагают, что эти мутации непосредственно усиливают образование вирусных частиц, в то время как в другой работе предполагается роль фосфорилирования белка N, позволяющая избежать ингибирования киназой GSK3 (ссылка [97]). Альтернативное объяснение состоит в том, что мутации R203K и G204R создают новый сайт регуляции транскрипции в середине гена N, что позволяет экспрессировать укороченную форму белка N (называемую «N*» или «N.iORF3»), которая может повышать инфекционность вируса за счет усиления антагонизма по отношению к интерферону [98, 99]. Интересно, что существует еще несколько примеров линий SARS-CoV-2, в которых появляются новые последовательности сайтов регуляции транскрипции, что может приводить к экспрессии укороченных белковых продуктов либо внутри, либо вне рамки считывания; это особенно выражено в случае неструктурного белка 16 (NSP16) [98].
Наряду с N-белком, мутации в M- и Е-белках также участвуют в модуляции инфекционной способности SARS-CoV-2. Было показано, что замены в М- и Е-белках BA.1 («омикрон») ослабляют проникновение вирусоподобных частиц в клетки, хотя эти мутации компенсируются дальнейшими заменами в S- и N-белках [100]. Е-белки коронавируса выполняют несколько функций, одна из которых заключается в том, чтобы действовать как катионный канал — вероятно, в компартментах эндоплазматического ретикулума (ЭПР) и аппарата Гольджи для регуляции нескольких стадий жизненного цикла вируса [101]. Было показано, что мутация T9I, обнаруженная в Е-белке «омикрона», ослабляет активность этого ионного канала in vitro [102], хотя ее функциональные последствия неясны.
Хотя ORF1ab составляет две трети генома SARS-CoV-2, он остается областью, где влияние вариантных мутаций наименее изучено. Единственным исключением является делеция в позициях 106–108 в NSP6 — консервативная мутация для всех ВВО, кроме «дельты». NSP6 представляет собой белок, пронизывающий мембрану несколько раз и связанный с формированием органеллы для репликации коронавируса — мембранной структуры, образующейся из ЭПР, которая формируется во время инфицирования, обеспечивая компартмент для репликации вирусной РНК, защищенный от врожденного иммунитета [103]. Недавнее исследование показало, что NSP6 образует гомодимеры и опосредует формирование «застегнутого» ЭПР — узких и ограниченных мембранных каналов, которые соединяют «двойные мембранные везикулы», служащие основным местом репликации вирусного генома [103]. Было обнаружено, что делеция 106–108 в NSP6 специфически усиливает образование такого «застегнутого» ЭПР, что указывает на потенциальную специфичную к хозяину адаптацию. Точный механизм такого усиления еще предстоит выяснить, хотя авторы исследования предположили, что делеция устраняет предполагаемый сайт О-гликозилирования. В настоящее время остается неясным, почему эта адаптация никогда не наблюдалась у «дельты» и ряда других вариантов, несмотря на легкость, с которой может произойти эта делеция.
Данные об адаптивных изменениях в дополнительных белках, помимо структурных белков и белков ORF1ab, которые рассматриваются далее в этом обзоре, очень ограничены. К сожалению, экспериментальная характеристика неспайковых мутаций и любых связанных с ними адаптаций к людям в вариантах SARS-CoV-2 сильно отстает от таковой для Spike-белка. Это связано с рядом факторов, в том числе с повсеместным распространением псевдовирусной технологии, используемой для изучения фенотипов Spike-белка на фоне технической сложности обратной генетики (обычно необходимой для вирусологических исследований мутаций вне Spike-белка) и нехваткой систем in vitro для исследования неспайковых белков. Из текущей работы ясно, что неспайковые адаптации вносят большой вклад в приспособленность и патогенность вируса, и для современных исследований жизненно важно продолжать развитие систем для изучения этих областей.
T-клеточный иммунный ответ и антигенное избегание
Т-клетки — основная часть адаптивного иммунного ответа на инфицирование SARS-CoV-2, при этом выраженный CD4+ Т-клеточный и CD8+ Т-клеточный ответ наблюдается у большинства людей. Несколько исследований указывают на важную роль Т-клеточного иммунитета в защите от тяжелой формы COVID-19, хотя эта роль сложнее и содержит больше нюансов, нежели хорошо охарактеризованная связь защиты от инфекции и ответа с участием нейтрализующих антител [105, 106]. Хотя ответы Т-клеток-хелперов CD4+, вероятно, в целом важны для образования антител, роль Т-клеток в снижении тяжести заболевания может быть относительно более важной в сценариях, когда ответы с участием нейтрализующих антител снижены или еще не обнаруживаются. Хотя ответы Т-клеток-хелперов CD4+, вероятно, в целом имеют большое значение для образования антител, роль Т-клеток в снижении тяжести заболевания может быть относительно более важной в сценариях, когда ответы нейтрализующих антител снижены или еще не обнаруживаются. Ранняя индукция Т-лимфоцитов, специфичных к SARS-CoV-2, чаще наблюдается при легких инфекциях, чем при тяжелых [107], а Т-клетки CD8+ могут быть особенно важны для облегчения тяжелых исходов у пациентов с дефицитом В-клеток [108]. Более того, полностью функциональный CD8+ Т-клеточный ответ активируется через одну неделю после введения первой дозы мРНК-вакцины BNT162b2 (Pfizer-BioNTech) — в период, когда нейтрализующие антитела индуцируются не полностью; это говорит в пользу того, что ранняя индуцированная вакцинами защита может в основном зависеть от Т-клеток [109].
Учитывая неотъемлемую роль Т-клеток в иммунитете против SARS-CoV-2, существует потенциальная возможность давления отбора, которое приведет вирус к избеганию Т-клеток, хотя степень воздействия мутаций SARS-CoV-2 на Т-клетки в настоящее время изучена плохо. Функциональные реакции Т-клеток направлены против нескольких вирусных белков, при этом величина ответа коррелирует с уровнями экспрессии этих белков. Преобладают реакции на Spike-белок, N-белок и М-белок, заметные реакции также наблюдаются на ORF3a и неструктурные белки NSP3 и NSP12 [110]. Поскольку T-клеточный ответ направлен на эпитопы в геноме SARS-CoV-2, признаки избегания Т-клеточного ответа распределены шире, нежели изменения, вызванные антителами, которые сконцентрированы в преобладающих эпитопах Spike-белка (рис. 3). В нескольких исследованиях отмечена эволюция внутри хозяина в эпитопах Т-клеток, что может служить прямым доказательством ускользания от Т-клеточного иммунитета. В одном исследовании отмечены мутации в CD8+-эпитопах N-белка (M322I и L331F), M-белка (L90F) и Spike-белка (L270F) в составе минорных вариантов в ходе острых инфекций, результатом которых стала утрата эпитопспецифичных реакций [111]. Длительная инфекция SARS-CoV-2 у хозяев с ослабленным иммунитетом может создавать более благоприятные условия для избегания Т-клеток — подобно тому, как это происходит при подробно изученном заражении ВИЧ [112]. Возникновение мутации NSP3 T504P, приводящей к потере эпитопного ответа CD8+, было отмечено у нескольких лиц с нарушением гуморального иммунитета, но с сохранением Т-клеточного ответа в контексте хронической инфекции SARS-CoV-2 [113, 114]. Эти результаты ограничены несколькими случаями, что говорит о необходимости более проспективных когортных исследований, систематически оценивающих риск избегания вирусом Т-клеток в определенных популяциях пациентов.
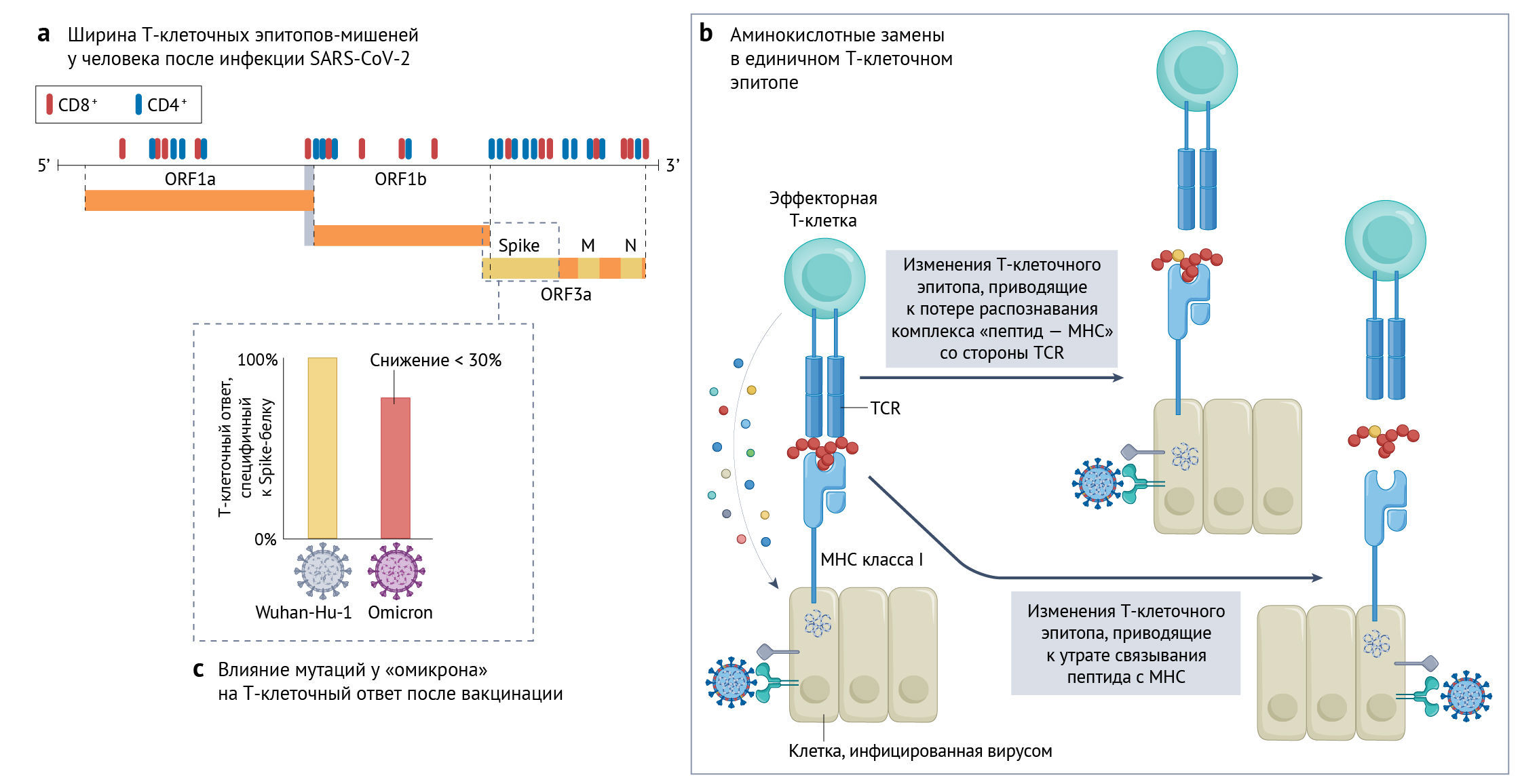
b. пример того, как замена аминокислоты в одном эпитопе может влиять на специфические к эпитопу ответы цитотоксических Т-клеток, тем самым ингибируя элиминацию инфицированных вирусом клеток [115]. Было показано, что уклонение SARS-CoV-2 от Т-клеток является следствием нарушения связывания пептида с главным комплексом гистосовместимости (MHC) или плохого связывания Т-клеточного рецептора (TCR) с комплексом «пептид–MHC».
c. Хотя Т-клеточный ответ на вакцинацию направлен только на Spike-белок, даже множественные мутации Spike-белка в рассматриваемом варианте «омикрон» снижают индуцированный вакцинами Т-клеточный ответ, специфичный к Spike-белку, менее чем на 30% со значительной межиндивидуальной вариабельностью [120, 191]. M — мембранный белок; N — белок нуклеокапсида.
Несколько мутаций в иммунодоминантных для CD8+ T-клеток эпитопах ORF3A и N-белка, которые приводят к полной потере распознавания, возникли независимо у множества линий SARS-COV-2 [115]. Среди них — P13L в N-белке, присутствующая у «омикрона» в вырезанном эпитопе CD8+ B*27:05. Учитывая гипотезу о том, что ВВО возникают при хронических инфекциях, можно предположить, что наличие р13L у ВВО «омикрон» отражает отбор под давлением Т-клеток во время такой хронической инфекции — в дополнение к набору мутаций в Spike-белке, возникших под давлением антител. L452R, обнаруженная в Spike- белке у вариантов «дельта», «эпсилон», «каппа» и BA.4/BA.5 приводит к потере A*24:02-CD8+-ответа [116]. Замена P272L в Spike-белке возникла во многих линиях по всему миру и приводит к потере доминантного эпитопа CD8+ HLA A*02:01 [117]. Роль Т-клеток в отборе, способствующем появлению этих изменений, в дополнение к избеганию антител и повышенному сродству к АПФ2, остается неясной. Другие мутации в Spike-белке ВВО, которые были связаны с потерей специфических ответов CD4+, включают L18F, D80A и D215G у варианта «бета» и D1118H — у варианта «альфа» [118, 119]. В настоящее время неизвестно, в какой степени эти наблюдения также отражают случайное влияние мутаций, движущей силой которых служит давление отбора по другим параметрам, на реакции T-клеток.
Несмотря на потерю этих специфических ответов, несколько исследований показывают, что общий Т-клеточный ответ, вызванный инфекциями и вакцинами первого поколения, сохраняется в отношении большинства ВВО [105, 118–120]. Даже обширные мутации в Spike-белке «омикрона» приводят лишь к незначительному — менее чем на 30% — снижению общего ответа клеток CD4+ и CD8+ со значительной вариабельностью между индивидуумами [119, 120]. Большинство происходящих с высокой частотой ответов CD4+ на эпитоп в Spike-белке сосредоточены на дискретном регионе N-концевого домена, С-конце и области слияния; при этом в RBD их очень мало [110]. Явных «горячих точек» для эпитопов CD8+ Spike-белка нет [111]. Следовательно, мутации, сосредоточенные в RBD и на N-конце Spike-белка, которые обнаружены у многих ВВО и возникли, как считается, для избегания антител и повышения аффинности к АПФ2, могут иметь ограниченное влияние на общий Т-клеточный ответ. Таким образом, большинство T-клеточных эпитопов консервативны в различных ВВО; и это, по-видимому, вносит вклад в сохранение эффективности вакцин для защиты от госпитализации и летального исхода при заражении «омикроном» после второй и третьей доз по сравнению с отсутствием вакцинации [112].
Другой ключевой причиной умеренного влияния вариантов на Т-клеточный иммунитет является широта генерируемого ответа, при этом каждый отдельный ответ после заражения нарастает на 30–40 эпитопов после [110]. На популяционном уровне также существует гораздо большая гетерогенность ответа Т-клеток по сравнению с антительными реакциями из-за ряда полиморфизмов, присутствующих в генах HLA человека. Однако на уровне индивидуумов примерно у 15% выздоравливающих и вакцинированных доноров было зарегистрировано значительное снижение специфичных к Spike-белку ответов CD8+ на «омикрон» [117]; при этом в другом исследовании отмечается потеря более чем 50% ответов CD4+ Т-клеток и CD8+ Т-клеток у ~ 20% протестированных лиц [122]. Хотя возможность обобщать эти результаты ограничена небольшими размерами выборки и распределением HLA в исследуемых популяциях, тем не менее, они подчеркивают потенциальное влияние ВВО на Т-клеточный ответ у некоторых людей, у которых иммунитет к Spike-белку возникает только в результате вакцинации.
Представляется вероятным, что уклонение от антител и повышенная трансмиссивность будут продолжать оставаться более мощными движущими силами для возникающих ВВО, чем значительное избегание Т-клеток. Сложно предсказать, будет ли со временем происходить медленная и последовательная потеря эпитопов CD8+ — подобно длительной адаптации гриппа H3N2 [97]. Избегание T-клеток может происходить с помощью нескольких механизмов [98]. Замены аминокислот внутри эпитопов или в примыкающих областях могут нарушать процессинг антигена, а замены якорных остатков могут мешать связыванию эпитопов с главными комплексами гистосовместимости (MHC) [123]. Оба этих механизма могут привести к необратимой потере способности Т-клеток отвечать на конкретный эпитоп. Напротив, результатом изменений, которые нарушают связывание T-клеточного рецептора с комплексом «пептид — MHC», может стать частичное или полное избегание. Последний сценарий можно легко предотвратить с помощью T-клеточных ответов de novo, используя альтернативные репертуары T-клеточных рецепторов, как было описано ранее для ВИЧ1-инфекции [124].
Помимо потенциального избегания Т-клеток, SARS-CoV-2, как и многие другие вирусы, напрямую подавляет экспрессию MHC I класса (MHC-I) в инфицированных клетках, чтобы избежать распознавания Т-клетками. Лучше всего это описано для вспомогательного белка ORF8 [125, 126]. Кроме того, сообщалось, что экспрессию MHC-I также подавляет ORF7a (а также ORF3a и ORF6 [127, 128]), хотя неясно, является ли это специфическим эффектом или просто результатом неспецифической фрагментации аппарата Гольджи [130]. В недавней работе было показано, что часто встречающиеся у ВВО замены не влияют на способность ORF8 подавлять экспрессию MHC-I — за исключением преждевременного стоп-кодона в 27 аминокислоте у варианта «альфа», в результате чего экспрессируется укороченная, нефункциональная форма ORF8 [126, 131]. Несмотря на укорочение ORF8, в контексте заражения «альфа» все еще подавляет экспрессию MHC-I; это подразумевает, что вариант «альфа» или, возможно, SARS-CoV-2 в целом развил дополнительные механизмы для ингибирования этого пути.
Врожденный иммунитет и варианты SARS-CoV-2
Врожденный иммунитет — неотъемлемая часть защиты хозяина от патогенов, которая играет ключевую роль в раннем контроле вирусов и регуляции адаптивных иммунных реакций [132]. Врожденный иммунный ответ особенно важен в случае новых вирусов, таких как зоонозные патогены, для которых обычно нет предсуществующего адаптивного иммунитета. Становится удивительным, когда новые зоонозные вирусы успешно распространяются среди людей благодаря эффективному противодействию врожденным защитным системам хозяина, несмотря на недавнюю успешную репликацию и передачу в неродственных видах-хозяевах с их типично несходной врожденной иммунной системой. Следствием надежности врожденной иммунной системы человека является то, что доля зоонозных вирусов, вызывающих пандемии, чрезвычайно низка, а свойства, развившиеся у вируса внутри видов-резервуаров SARS-CoV-2 (в частности, универсальная тропность к хозяину, включая людей), в сочетании с приобретением FCS Spike-белком (возможно, на уровне промежуточных видов-хозяев) были критическими особенностями для начала пандемии COVID-19. Примечательно, что родственный вирус атипичной пневмонии SARS-CoV не прижился в популяции людей, несмотря на то, что он также обладает высокой трансмиссивностью, а его исчезновение объясняется меньшей склонностью к бессимптомному распространению по сравнению с SARS-CoV-2, что упрощает выявление случаев заражения. Важно отметить, что с появлением ВВО SARS-CoV-2 стало ясно, что они приобретают адаптации для более эффективного заражения людей, и отчасти это происходит за счет усиленного уклонения от врожденного иммунитета. Действительно, у «альфы» [133, 134] и более поздних ВВО [77, 135, 136] развилась пониженная чувствительность к интерферонам, и это является ключевым фактором давления отбора при передаче вируса в человеческой популяции. Удивительно, но вместо того, чтобы адаптироваться для специфического противодействия человеческим белкам, подлинии вариантов «альфа» и «омикрон» BA.4 и BA.5 частично достигли этого за счет усиления экспрессии вирусных белков, подавляющих врожденный иммунитет, — особенно белка ORF6 [77]. ORF6 ингибирует ядерный транспорт транскрипционных факторов STAT1 и IRF3 [137], которые контролируют экспрессию противовирусных белков и растворимых провоспалительных медиаторов. У «альфы» также усилилась экспрессия ORF9b (который ингибирует сигнальные пути после узнавания РНК) и N-белка (который секвестирует вирусную РНК, чтобы предотвратить запуск механизмов ее узнавания [98]), а также экспрессия de novo N*/N.iORF3 — укороченной с N-конца формы N-белка, которая проявляет некоторый антагонизм к интерферонам, но экспрессируется на низких уровнях [98]. Повышенные уровни этих белков, вероятно, являются результатом мутаций в регуляторных областях, которые модулируют субгеномный синтез РНК и экспрессию белка. Это подчеркивает важность внеспайковых изменений в определении свойств ВВО, особенно в регуляторных областях. Важно, что мутации в последовательности Kozak N-белка, для которых ожидалось влияние на экспрессию N-белка у варианта «альфа» и перекрывающегося белка ORF9b, также появляются в доминирующих ВВО «дельта» и «омикрон», но их полное влияние на антагонизм этих ВВО по отношению к врожденному иммунитету еще предстоит определить. Взаимодействия между SARS-COV-2 и врожденным иммунитетом очень сложны. Например, вирусный антагонист ORF9b, по-видимому, негативно регулируется путем фосфорилирования киназами хозяина; это позволяет предположить, что его способность ингибировать врожденный иммунитет может выключаться на определенном этапе инфекции: возможно, когда в зараженных клетках запускаются реакции хозяина [133]. Такой механизм воспалительного переключения, который по существу регулирует реакцию хозяина на инфекцию, может стимулировать симптомологию и последующее распространение вируса путем изменения клеточной активации.
На сегодняшний день предполагается, что по крайней мере 15 белков SARS-CoV-2 участвуют в противодействии врожденным иммунным ответам (детально описано в [139, 140]). Эти белки обычно определяются с помощью репортерных скринингов, в которых вирусный белок интереса экспрессируется при моделируемом in vitro врожденном иммунном ответе, и оценивается способность этого белка противодействовать такому ответу. Обычно такие эксперименты не дают понимания механизмов, но они эффективны для обнаружения новых функций белков. У нескольких ВВО имеются аминокислотные замены во многих белках, участвующих в противодействии системе врожденного иммунитета, включая NSP1, NSP3, NSP6, ORF3a, ORF6, ORF7b, ORF8 и N-белок. Отражают ли эти кодирующие мутации адаптацию для лучшего противостояния врожденному иммунитету человека, в большинстве случаев еще предстоит установить. Более того, вклад этих белков в фенотип ВВО изучен плохо, так как для этого требуется трудоемкое создание изогенных мутантов с применением обратной генетики и оценка их влияния на репликацию, выработку интерферона и чувствительность.
Помимо снижения индукции интерферона, ВВО «альфа» и «омикрон» более устойчивы к к его ингибиторным противовирусным эффектам [126, 133–135, 141]. Лучше всего это описывается в связи с адаптациями со стороны Spike-белка, которые снижают чувствительность к индуцированным интерфероном трансмембранным (IFITM) рестрикционным факторам [76, 134, 142]. Точная роль белков IFITM во время репликации SARS-CoV-2 является спорным вопросом, при этом некоторые исследования показывают, что белки IFITM ингибируют проникновение SARS-CoV-2 в клетки — как это происходит в случае гриппа [19, 76, 134, 142], тогда как другие предполагают, что в некоторых контекстах они усиливают инфекцию способом, аналогичным описанному для OC43 и HKU1 [76, 134, 143, 144, 145]. Функция белков IFITM, вероятно, зависит от контекста, такого как конкретный путь проникновения, используемый конкретным вирусом в данном типе клеток или клеточной линии, использование живого вируса или псевдовируса и уровень экспрессии белка IFITM. Члены семейства IFITM представляют собой малые, стимулируемые интерферонами трансмембранные белки, связанные с различными клеточными мембранами; у человека IFITM1 обычно ассоциированы с мембранами клеточной поверхности, тогда как IFITM2 и IFITM3 преимущественно локализованы в поздних и ранних эндосомах, соответственно [76, 146]. Точный механизм, с помощью которого белки IFITM влияют на проникновение вируса, полностью не выяснен, но считается, что в нем участвует ингибирование слияния вирусных гликопротеинов с мембранами хозяина [147, 148]. Показано, что несколько ВВО обладают разной степенью чувствительности к ингибированию или активации белка IFITM, что довольно часто связано со специфическими путями проникновения или фенотипами расщепления фурином. Например, «омикрон», который более эффективно проникает в эндосомы, чем ранние варианты и другие ВВО, по-видимому, демонстрирует большее ингибирование (или в некоторых случаях усиление) эндосомными белками IFITM, хотя это, по-видимому, сильно зависит от используемой клеточной системы [32, 76, 145]. Возможно, это связано с тем, что «омикрон» либо не избегает адаптивного иммунитета (особенно в контексте индуцированных вакциной антител к Spike-белку), либо вынужден адаптироваться к использованию белков IFITM в качестве кофакторов для проникновения.
Пока неясно, как усиленное противодействие врожденному иммунитету может влиять на передачу вируса. Авторы предполагают, что способность SARS-CoV-2 к эффективному распространению тесно связана с его способностью уклоняться от врожденного иммунного ответа в первых клетках, которые сталкиваются с вирусом в дыхательных путях, и противодействовать ему. Действительно, ожидается, что эффективность инфекционных событий будет влиять на распространение вируса через дыхательные пути и, следовательно, на вероятность возникновения продуктивной инфекции в целом. Было показано, что для определения эффективности инфекции и исхода в случае других вирусов важен ответ интерферона типа I; это хорошо описано для лентивирусной инфекции макак [149].
Наконец, тот факт, что в настоящее время каждый ВВО эволюционировал независимо от предкового вируса, циркулировавшего в начале пандемии, означает, что каждый ВВО прошел свой мутационный путь, чтобы приобрести различные приспособления к организму человека. Как следствие, ВВО или даже другие варианты SARS-CoV-2 потенциально могут рекомбинировать, чтобы объединить независимо кодируемые адаптации и, следовательно, фенотипические преимущества различных вариантов геномов (вставка 2).
Антигенное дистанцирование в определении передачи и приспособленности
Постоянно меняющийся популяционный иммунитет создает для вариантов вируса динамический ландшафт приспособленности, поскольку эта приспособленность в такой же степени зависит от приобретенного иммунитета, как и от набора уникальных мутаций вируса. Хотя такие сложности, как широта и продолжительность иммунитета, являются важными факторами [150, 151], увеличение числа людей, подвергшихся воздействию SARS-CoV-2 в результате инфекции и/или вакцинации, приводит к тому, что популяция становится гораздо менее восприимчивой к большинству циркулирующих (и прошлых) вариантов, а число иммунологически наивных, полностью восприимчивых хозяев постоянно снижается (рис. 4).
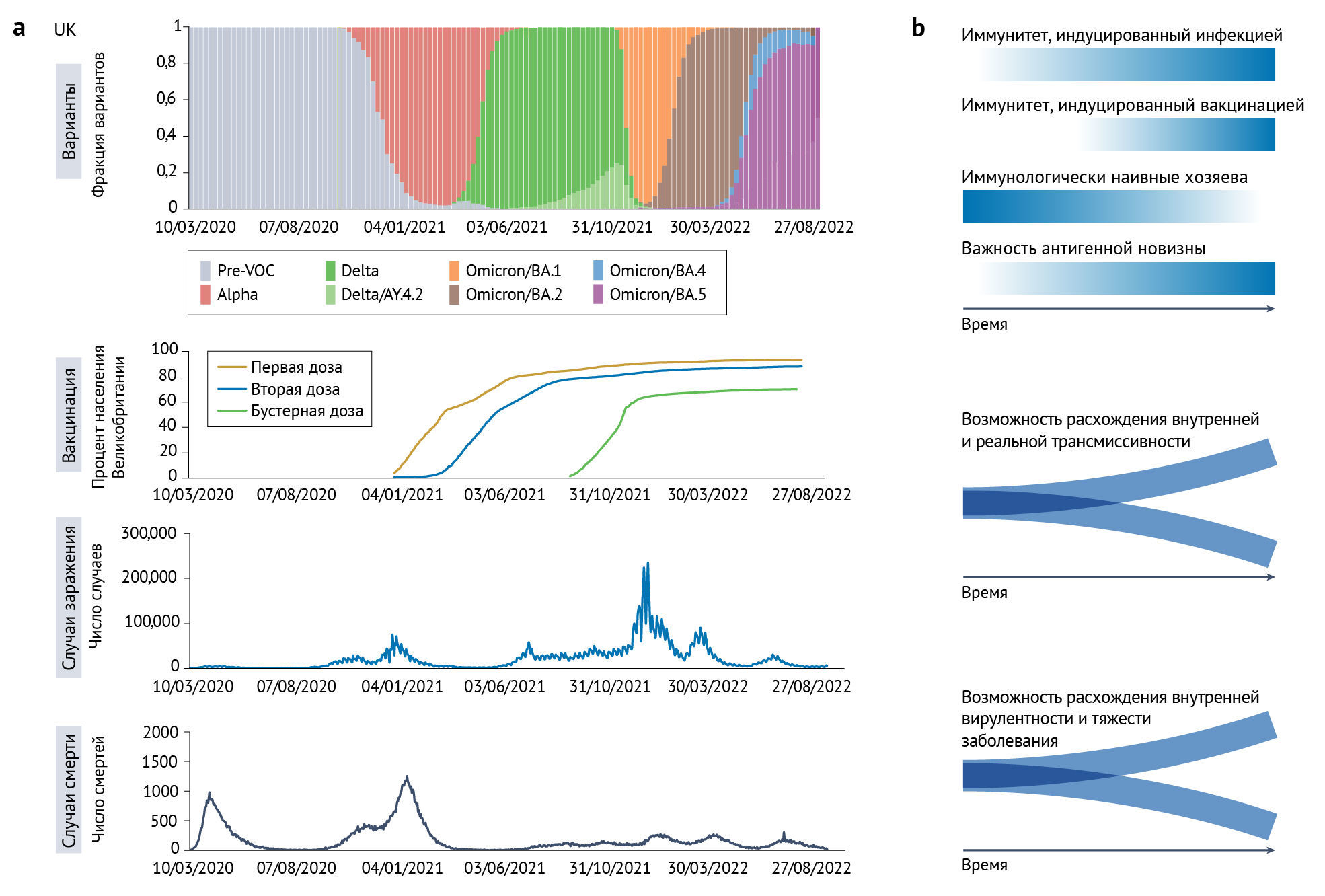
b. Схематическая визуализация динамической взаимосвязи между трансмиссивностью, антигенностью, вирулентностью и приспособленностью вариантов. По мере того, как популяционный иммунитет, возникающий в результате инфекции и вакцинации, увеличивается, доля полностью иммунологически наивных хозяев снижается (градиентные синие линии). Следовательно, важность антигенной новизны в определении приспособленности вариантов возрастает. Антигенное расстояние до ранее циркулирующих вариантов становится все более важным фактором, определяющим трансмиссивность вариантов; это увеличивает вероятность расхождения внутренней и реальной трансмиссивности. . Сходным образом, антигенное расстояние влияет на способность варианта заражать иммунных хозяев и вызывать у них заболевание, увеличивая вероятность отклонения внутренней вирулентности варианта от его клинического влияния в реальном мире.
В начале пандемии COVID-19, когда доля наивных хозяев была наибольшей, антигенная новизна имела мало эволюционных преимуществ по сравнению с вариантами дикого типа. Вместо этого отбор благоприятствовал вариантам, способным максимизировать репродуктивный успех за счет приспособлений со стороны внутренних биологических особенностей, таких как конформационные изменения Spike-белка, вызванные D614G — определяющая мутация линии PANGO B.1 — или фенотип с усилением расщепления фурином в сочетании с повышенным связыванием AПФ2, как у варианта «альфа» [27, 29]. По мере повышения иммунитета в популяции хозяина антигенная новизна варианта играла все более важную роль в его репродуктивном успехе по сравнению с внутренними биологическими изменениями [152]. Впоследствии ВВО «дельта» стал доминирующим во всем мире, вытеснив предыдущие варианты в странах с частично иммунными популяциями и средним или высоким охватом вакцинацией [23, 153]. Данные по нейтрализации вируса указывают на умеренное избегание нейтрализующих антител в случае ВВО «дельта» [23, 37–39], а данные об эффективности вакцин говорят о том, что антигенная новизна не была основной движущей силой повышения трансмиссивности [40–42, 154, 155]. Это свидетельствует в пользу того, что высокая приспособленность «дельты» в большей степени является следствием внутренних свойств вируса, таких как оптимизация расщепления фурином Spike-белка [22, 23, 69].
«Омикрон» демонстрирует беспрецедентную степень антигенной новизны по сравнению с другими вариантами [31, 33, 34], возможно, сравнимую с таковой для антигенного сдвига в случае гриппа [51]. Здесь «сдвиг» — накопление мутаций, участвующих в антигенном дистанцировании, возможно, по крайней мере отчасти возникает в контексте хронической инфекции (или инфекций) (вставка 1). Сравнение динамики передачи внутри вакцинированных и невакцинированных семей указывает на то, что ключевым фактором повышенной трансмиссивности «омикрона» (BA.1) по сравнению с «дельтой» в период их совместной циркуляции было избегание иммунной системы [86]. Показано, что подлиния «омикрона» BA.2 обладает даже большей способностью заражать как вакцинированных, так и невакцинированных людей, возможно, благодаря свойствам иммунного избегания, сходным с таковыми для BA.1, но с более высокой внутренней трансмиссивностью [156]. В последнее время BA.4, BA.5, BA.2.75 и их подлинии продемонстрировали не только лучшую способность избегать иммунитета по сравнению с вариантами, предшествовавшими «омикрону»; они могут избегать и иммунного ответа после заражения «омикроном», особенно BA.1 [46–49, 157–159]. Это в большой степени приписывается к мутациям в антигенно значимых положениях RBD; например, у BA.4/BA.5 это L452R и F486V [160]. Хотя потенциал для дальнейшей оптимизации передачи SARS-CoV-2 в человеческой популяции может оставаться, в настоящее время представляется вероятным, что антигенная новизна появляющихся вариантов и их способность избегать иммунитета будут главными факторами, определяющими приспособленность варианта и эволюционный успех в будущем. Следовательно, основным приоритетом исследований будет понимание сложности перекрестной защиты между вариантами.
Относительная тяжесть инфекции при разных вариантах SARS-CoV-2
Крайне важно понять, как вирулентность вариантов SARS-CoV-2 может развиваться в ответ на меняющееся давление отбора. Вирулентность патогена, наряду с иммунитетом, индивидуальной восприимчивостью, предрасположенностью к заболеванию и другими факторами со стороны хозяина вносит основной вклад в тяжесть заболевания и определяется в эволюционной литературе как повышенная заболеваемость и смертность людей вследствие инфекции. Вирулентность не обязательно снижается со временем в популяции хозяина; скорее, данные моделирования обычно показывают компромисс между скоростью передачи и вирулентностью [161, 162]. Однако предсказуемость эволюции вирулентности осложняется несколькими механизмами, включая конкуренцию внутри хозяина, изменение путей передачи и тропизма, а также взаимодействие с иммунной системой [162]. Например, неоднократные волны эпидемии, характерные для патогена с эволюцией антигенных свойств, могут приводить к отбору на высокую вирулентность патогена [163]. Оценка относительной вирулентности вариантов SARS-CoV-2 по тяжести заболевания у людей — сложная задача из-за изменения иммунного статуса и развития медицинских подходов на протяжении пандемии, хотя можно сравнивать тяжесть заболевания для вариантов, которые заразили одну и ту же популяцию в пределах данного периода [164]. Этот подход предполагает несоответствие в направлении изменения тяжести заболевания в ряду последовательно доминирующих вариантов SARS-CoV-2: успешные варианты продемонстрировали повышенную тяжесть заболевания, когда вариант «альфа» заменил B.1.177, а «дельта» заменил «альфу», что коррелирует с относительными изменениями трансмиссивности [164]. Напротив, в случае «омикрона» в период, когда он сосуществовал с «дельтой», тяжесть заболевания снижалась; такое снижение, видимо, отражает сложную комбинацию факторов, включая как более высокую скорость заражения у лиц с определенной степенью предшествующей инфекции, так и собственную низкую вирулентность [164–168]. Среди предложенных объяснений уменьшения тяжести заболевания — пониженная фузогенность Spike-белка, приводящая к меньшему повреждению тканей, и измененный тропизм, ограничивающийся в большей степени верхними дыхательными путями (из-за измененной роли TMPRSS2) [32, 75, 78, 79]. В этих предыдущих исследованиях сравнивали вирулентность вариантов, присутствующих в одной популяции, хотя возможности определить «внутреннюю» вирулентность непересекающихся вариантов, таких как «альфа» и «омикрон», которые циркулировали в популяциях с разным иммунным статусом, не было. Учитывая, что иммунный статус человека в дополнение к вероятности инфицирования влияет на тяжесть симптомов, антигенное «расстояние» между прошлым воздействием и дивергентным вариантом потенциально может способствовать возникновению заболевания у защищенного в других отношениях человека, что демонстрирует возможное расхождение между собственным опасным потенциалом вируса и фактической вирулентностью у инфицированных людей.
Дополнительный подход к оценке тяжести вариантов SARS-CoV-2 включает применение животных моделей (более подробно рассмотрено ранее [169]). Обычные модели на животных включали наивных грызунов, таких как трансгенные мыши, экспрессирующие человеческий AПФ2 под контролем промотора кератина 18 (обильно экспрессируемого в эпителиальных клетках [170]), или хомяков, чьи нативно экспрессируемые белки AПФ2 эффективно используются всеми текущими ВВО SARS-CoV-2 [32, 171]. Патогенность в таких моделях на грызунах чаще всего измеряют как функцию потери веса в процентах, иногда наряду с использованием кривых выживания и показателей легочной функции [172]. Модели на грызунах в значительной степени воспроизводят эквивалентные данные о серьезности эпидемиологических исследований на людях — например, «дельта» более патогенна, чем более ранние варианты, а «омикрон» менее патогенен, чем «дельта» [32, 69, 75, 173, 174]. Однако они имеют ряд ограничений. Примером могут служить недавние эпидемиологические данные из Гонконга, которые предполагают, что подлиния «омикрона» BA.2 вызывает заболевания, по тяжести сходные с таковыми для вариантов первой волны [175], в то время как модели на грызунах обычно показывают, что «омикрон» переносится легче, чем предыдущие варианты [32, 75, 173, 174, 176]. Это несоответствие потенциально может быть объяснено продолжающейся адаптацией SARS-CoV-2 к человеку в качестве хозяина, что приводит к сопутствующим отличиям адаптации у животных, использующихся для моделирования инфекции [32, 80, 177, 178].
Вставка 3. Приспособленность и антигенные свойства
Дарвиновская приспособленность [209], обычно называемая просто «приспособленностью», отличается от репликативной приспособленности — способности вируса производить инфекционное потомство, измеряемой экспериментально в культивируемых клетках, в культуре тканей или у отдельных хозяев [210, 211]. Напротив, более широкое определение приспособленности подразумевает репродуктивный успех, поэтому она сильно зависит от контекста, часто меняясь во времени и в разных локациях. Приспособленность данного варианта SARS-CoV-2 будет зависеть от изменяющегося иммунного профиля популяции хозяина, в которой он циркулирует, а успех отдельного варианта зависит от свойств конкурирующих вариантов в популяции вируса и процессов стохастического отбора. По мере того, как иммунитет в популяции хозяина возрастает, может произойти так, что вариант, ранее высокотрансмиссивный в неиммунной наивной популяции, теперь будет менее приспособленным по сравнению с более развитыми вариантами (рис. 4b). Хотя внутренняя трансмиссивность основана только на биологических свойствах вируса, фактическая трансмиссивность зависит от этих свойств в контексте популяционного иммунитета, включая иммунитета хозяина от предыдущих инфекций, антигенные свойства штамма и стохастические эффекты. Более того, хотя возможно, что варианты могут различаться по своей относительной антигенности — уровню иммунного ответа, который они стимулируют, — различия в степени антигенного сходства с другими вариантами обычно более значимы. По мере повышения иммунитета в популяции хозяев антигенное расстояние между текущим вариантом и ранее циркулировавшими вариантами может определять способность заражать хозяев, реплицироваться внутри них и передаваться между ними. Следовательно, эволюция антигенной новизны становится критическим фактором, определяющим репродуктивный успех и приспособленность вариантов.
Тяжесть заболевания после заражения SARS-CoV-2 коррелирует с несколькими факторами риска, включая пожилой возраст, принадлежность к мужскому полу и сопутствующие заболевания, такие как ожирение и иммунодефицит, а также ряд воспалительных маркеров [179]. Несколько недавних генетических исследований сосредоточились на характеристиках, которые могут объяснить, почему некоторые люди менее восприимчивы к инфекциям SARS-CoV-2, а у других развиваются более тяжелые симптомы [180]. Тем не менее, существует настоятельная необходимость объединить эти результаты с данными о конкретных вариантах и терапевтических воздействиях (то есть вакцинах, лекарствах и моноклональных антителах), которые могут непосредственно влиять на наблюдаемые фенотипы [179]. Кроме того, хотя наиболее распространенными и легко измеримыми исходами острых инфекций являются госпитализация или смерть, исходы, которые измерить труднее, — такие как первичная симптоматика [181] или «постковидный» синдром — также сильно различаются в зависимости от вариантов, и, вероятно, — по предшествующему иммунному статусу.
Заключение
SARS-CoV-2 три года циркулирует в человеческой популяции, заражая сотни миллионов человек. Тем не менее, он остается относительно новым для человека вирусом, который продолжает эволюционировать и приобретать адаптации к своему новому виду-хозяину. Уникальные наборы данных о последовательностях генома SARS-CoV-2, созданные во всем мире, показали появление полезных мутаций в режиме реального времени и помогли лучше понять внутренние свойства взаимодействий между вирусом и хозяином для лабораторных экспериментов. Несмотря на хорошее понимание биологии SARS-CoV-2, приспособленность вируса очень динамична, и способность SARS-CoV-2 заражать людей, размножаться внутри них и распространяться по популяции очевидно зависит от конкретного иммунного контекста в разные периоды пандемии. На сегодняшний день во всем мире доминирует «омикрон», а именно быстро возникшие его подлинии BA.2 и BA.5. Хотя знания о SARS-CoV-2 улучшаются, эволюция вируса по своей природе непредсказуема, и вероятным будущим сценарием является появление нового ВВО, который по своим антигенным свойствам и, потенциально, фенотипически отличается от ранних форм «омикрона». В то же время продолжает накапливаться популяционный иммунитет против SARS-CoV-2, что вполне может компенсировать появление в будущем варианта, вызывающего более тяжелое заболевание, и привести к более легкому протеканию острой инфекции.
Все предшественники ВВО развились из вирусного предка («пре-ВВО»), существовавшего во время первой волны пандемии, по различным, но часто конвергентным путям, направленным на более эффективное заражение и распространение среди людей и устойчивость к антителам, Т-клеточному и врожденному иммунитету. Необходимые для этого адаптации, как обсуждается в этом обзоре, представляют собой набор изменений в организме хозяина, обусловленных внутренними свойствами вируса и избеганием врожденного или адаптивного иммунитета (вставка 3). Согласно доминирующей гипотезе, новые варианты образуются в ходе хронических инфекций у лиц с ослабленным иммунитетом; в этом случае вирус способен вызвать персистирующую инфекцию вследствие нарушения иммунных функций [67, 114, 182]. Эта гипотеза объясняет скачкообразные изменения в ходе кажущейся быстрой эволюции, наблюдаемые перед появлением новых вариантов [67, 183, 184]. Так или иначе, следует отметить, что будущие варианты, возможно, произойдут от предшествующих или современных ВВО; в качестве примера можно привести недавний всплеск вариантов «омикрон» «второго поколения», возникших из BA.2, такие как BA.2.75, BJ.1 и BA.2.10.4 [185]. В то время как внутрилинейная рекомбинация дает вирусу возможность получить дополнительные адаптации и фенотипические преимущества от циркулирующих вариантов с отдаленным родством, до появления ВВО рекомбинанты оказывали лишь незначительное влияние на течение пандемии (вставка 2). Кроме того, несмотря на то, что в настоящее время данных об установлении долгосрочной циркуляции и эволюции у видов животных-резервуаров очень мало, необходим интенсивный и активный надзор за восприимчивыми видами, поскольку регистрируется обратный зооноз [4, 5, 186]. У многих стран мало возможностей для секвенирования; существуют также места, в которых ранее эпиднадзор был хорошим, а затем объемы секвенирования снизились или же оно было полностью прекращено. Это создает проблемы, поскольку отсутствие геномного надзора будет означать, что будущие варианты будут выявлены намного позже или смогут циркулировать на низких уровнях до возможного обнаружения. Таким образом, требуется широкомасштабный и соразмерный охват эпиднадзором для быстрого выявления потенциальных новых ВВО среди людей и сообществ до того, как новые штаммы распространятся более широко.
