
Движение и рефлексы

Аристотелю приписывается широко известная фраза: “Движение — это жизнь”. Сеченов в своей работе “Рефлексы головного мозга” высказывается о движении более поэтично: “Всё бесконечное разнообразие внешних проявлений мозговой деятельности сводится окончательно к одному лишь явлению — мышечному движению. Смеётся ли ребенок при виде игрушки, улыбается ли Гарибальди, когда его гонят за излишнюю любовь к Родине, дрожит ли девушка при первой мысли о любви, создаёт ли Ньютон мировые законы и пишет их на бумаге — везде окончательным фактом является мышечное движение”.
Чем же обеспечивается движение? Какие пути и структуры помогают нам не только перемещаться в пространстве, но и совершать более мелкие движения?
Все механизмы, участвующие в реализации движения, довольно многочисленны и сложны. Условно все движения разделяют на две большие группы (рефлекторные и произвольные), но как такового разделения между ними нет: в течение жизни многие произвольные движения, которые мы выполняем рутинно, начинают совершаться нами “на автомате”, что значительно уменьшает энергетические затраты и увеличивает эффективность.
Как известно, своеобразным управляющим пунктом в нашем организме является головной мозг. В норме он осуществляет контроль над всеми органами и системами, и мышцы не исключение. За проведение сигнала от анализатора к эффектору и осуществление осознанных движений отвечает корково-мышечный путь.
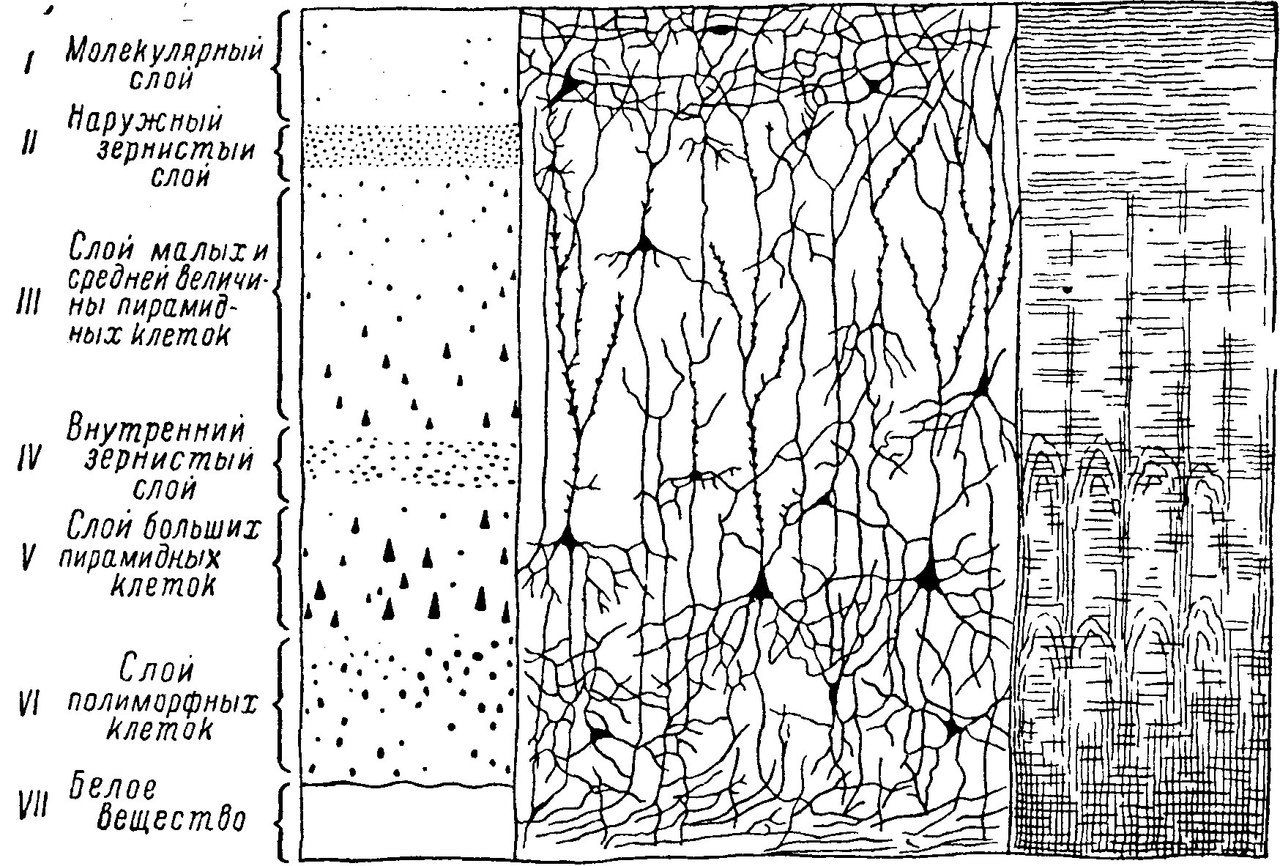
Ядра двигательного анализатора находятся в предцентральной извилине (ПИ), а также в задних отделах средней и нижней лобных извилин. Передача импульса осуществляется по волокнам пирамидных путей, берущих начало от клеток Беца (пятого слоя коры головного мозга).
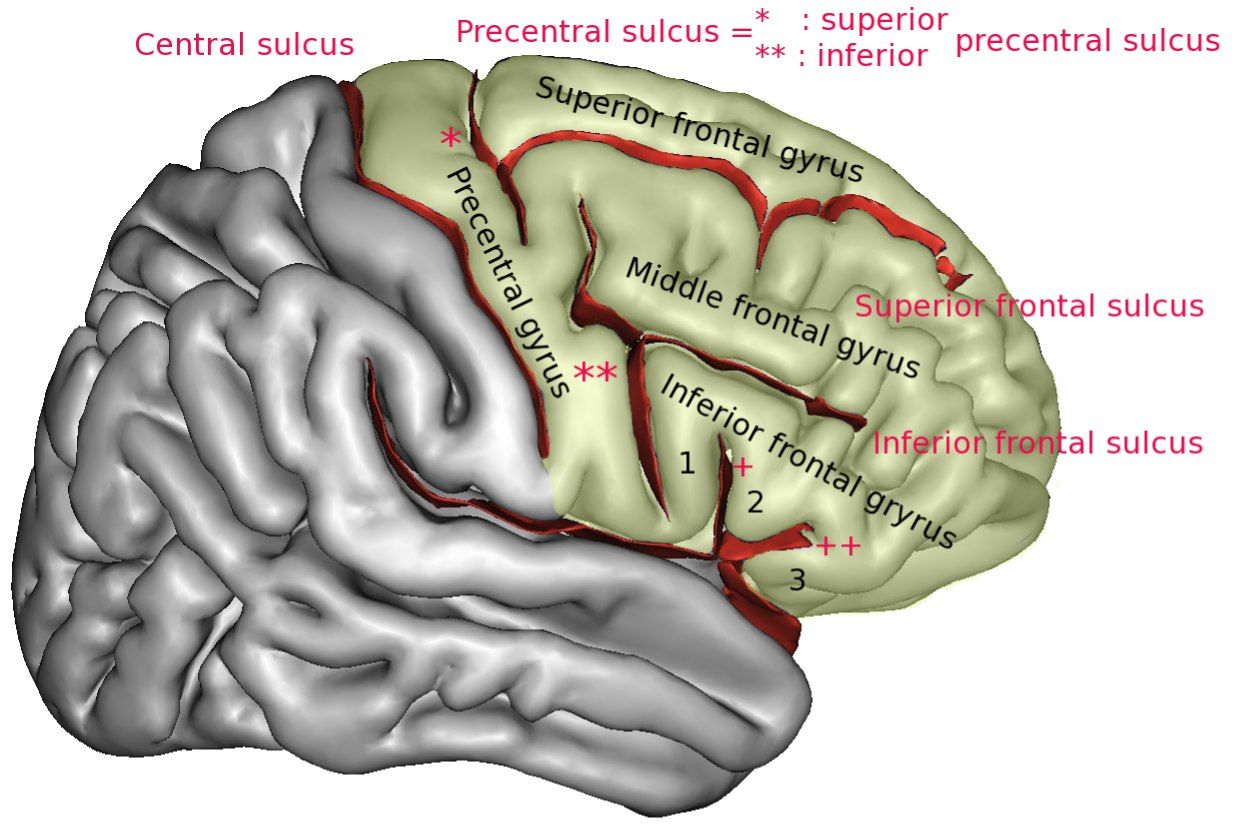
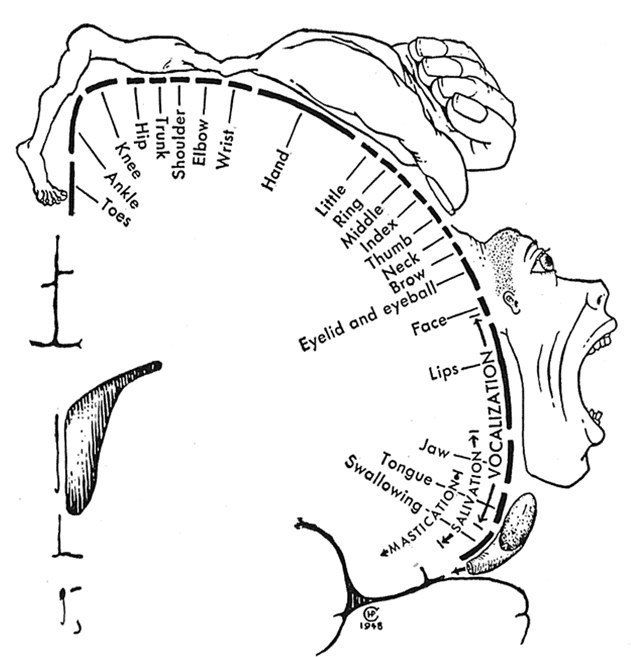
Зоны, иннервируемые клетками ПИ, традиционно изображаются на схемах в виде “моторного гомункулуса”: в верхней части ПИ и парацентральной дольке располагаются корковые отделы двигательных анализаторов (КОДА) мышц нижней конечности, в средней — туловища, верхней конечности, а также шеи, в нижней — мышц языка, лица и глотки. В заднем отделе средней лобной извилины расположен КОДА сочетанного поворота головы и глаз, там же находится двигательный анализатор письменной речи (написание букв, цифр и тд.). Задний отдел нижней лобной извилины является КОДА устной речи.
Начавшись в ПИ, пирамидный путь разделяется на tractus corticospinalis и tractus corticonuclearis.
Кортикоспинальный путь идёт через белое вещество больших полушарий, образуя при этом лучистый венец. Он направляется в передний отдел задней ножки внутренней капсулы. Далее, на границе продолговатого и спинного мозга, происходит частичный перекрест: 80% волокон переходит на другую сторону, образуя при этом боковой канатик, и спускается в составе tractus corticospinalis lateralis. Оставшиеся 20% идут ниже в составе переднего канатика и образуют tractus corticospinalis anterior, в котором они частично перекрещиваются.
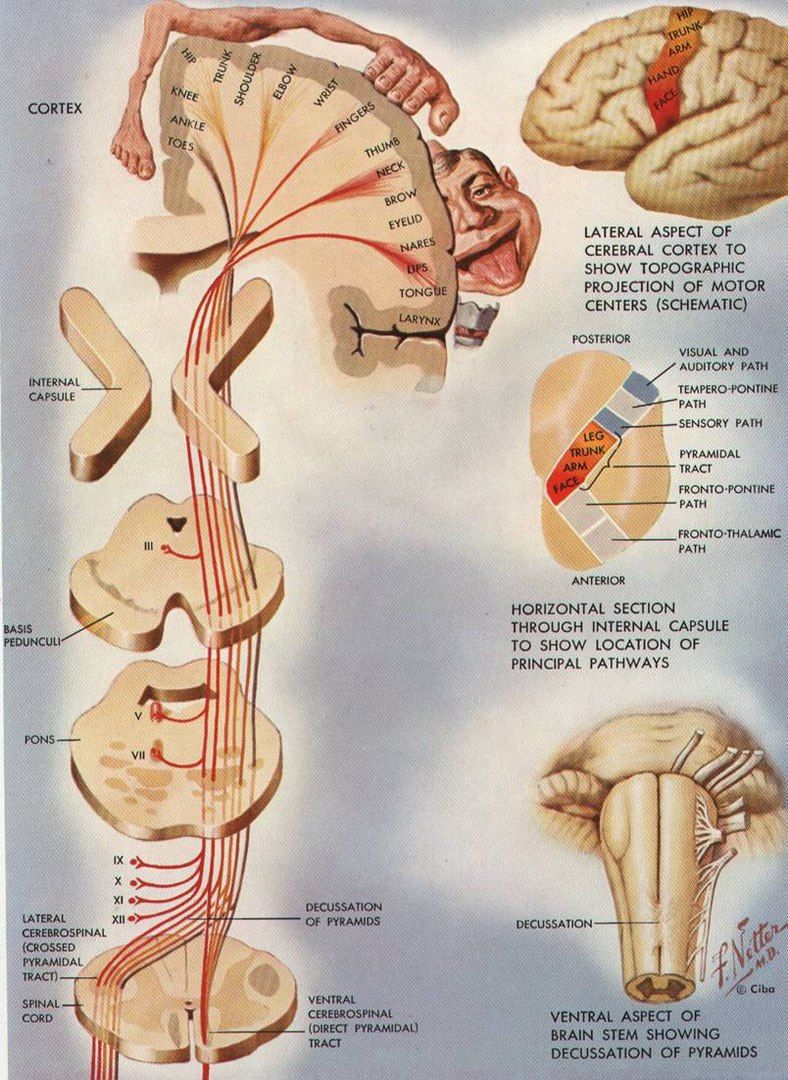
Смысл всех вышеописанных перекрестов заключается в обеспечении двусторонней иннервации туловища.
Кортиконуклеарный путь берёт начало от клеток нижнего отдела ПИ и проходит через колено внутренней капсулы. Около ядер черепных нервов происходит частичный надъядерный перекрест волокон, что обеспечивает двустороннюю иннервацию мышц лица, глотки и гортани (за исключением языка и нижней половины лица, которые иннервируются односторонне из-за полного перекреста волокон).
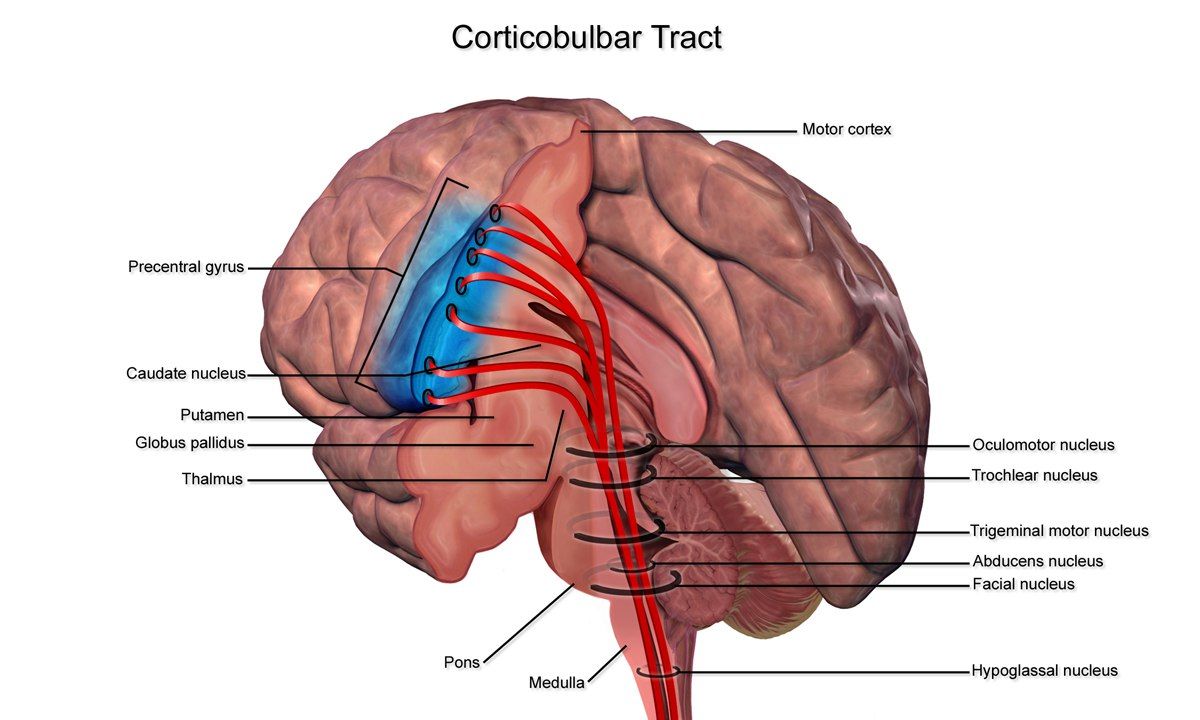
Далее происходит переход от центрального двигательного нейрона к периферическому. Периферический двигательный нейрон включает мотонейроны передних рогов спинного мозга, передние корешки, волокна в составе сплетений и периферических нервов.
Мотонейроны для мышц разных групп располагаются на уровне разных сегментов. Мотонейроны для мышц верхней конечности располагаются в шейном утолщении на уровне С5-Th1, нижней конечности — в пояснично-крестцовом утолщении на уровне L1-S2, а туловища — на уровне грудных сегментов.
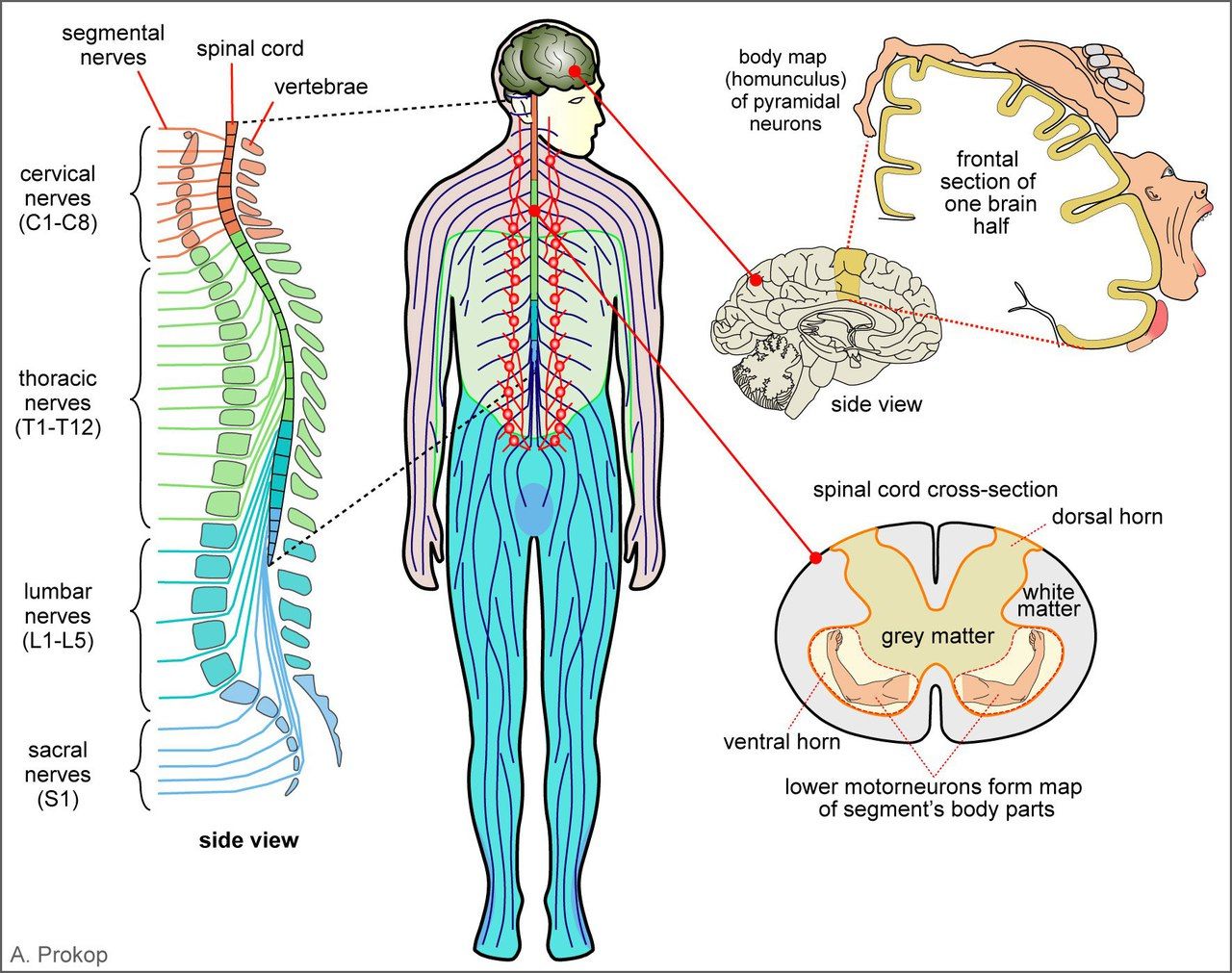
Покидая спинной мозг, аксоны передних рогов формируют передние корешки. После прохождения спинномозгового ганглия они присоединяются к задним корешкам, образуются смешанные нервы, которые идут к мышцам.
Совместно с центральным мотонейроном периферический обеспечивает нормальную выраженность сегментарных рефлексов, которые подразделяются на глубокие и поверхностные, а также на условные и безусловные.
Основой любого рефлекса считается рефлекторная дуга (закрепившийся термин, который, однако, потерял свою актуальность с открытием обратной афферентации, и слово “дуга” было заменено на “кольцо”).
Для осуществления рефлекса необходимо три составляющих: афферентное (рецептор), вставочное (связывают рецептор с эффектором) и эфферентное звено (нейрон с окончанием в ткани органа).
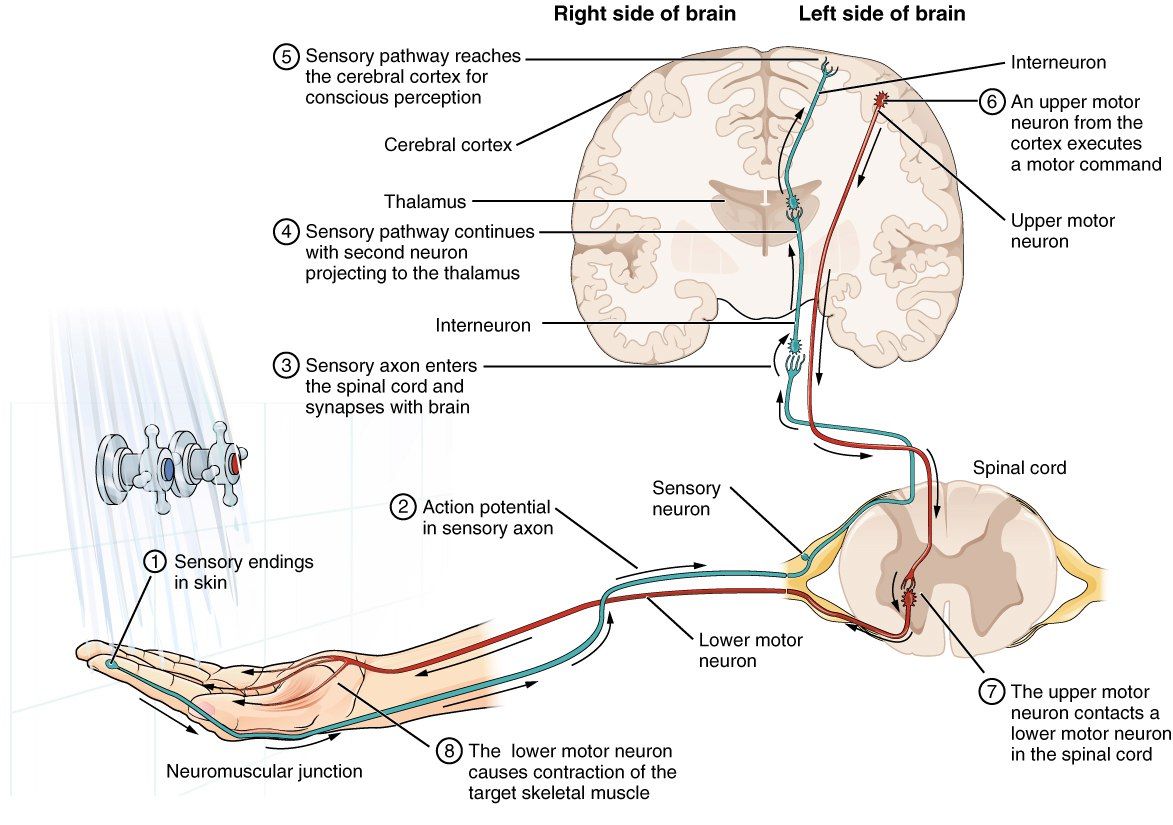 Мышечный рефлекс запускается при воздействии внешнего стимула на рецептор, расположенный в мышце. Однако, согласно современной концепции о рефлекторном кольце, возникает не один, а два потока импульсов, один из которых идёт к клеткам передних рогов соответствующих сегментов (иннервирующих агонисты и приводящих к их сокращению), а второй — к клеткам передних рогов соседних сегментов (иннервирующих мышцы антагонисты и приводящих к их расслаблению). Вставочные нейроны, через которые проходит второй поток импульсов, производят тормозные импульсы, из-за чего при сокращении агонистов и растяжении антагонистов последние не сокращаются и становится возможно осуществление рефлекса.
Мышечный рефлекс запускается при воздействии внешнего стимула на рецептор, расположенный в мышце. Однако, согласно современной концепции о рефлекторном кольце, возникает не один, а два потока импульсов, один из которых идёт к клеткам передних рогов соответствующих сегментов (иннервирующих агонисты и приводящих к их сокращению), а второй — к клеткам передних рогов соседних сегментов (иннервирующих мышцы антагонисты и приводящих к их расслаблению). Вставочные нейроны, через которые проходит второй поток импульсов, производят тормозные импульсы, из-за чего при сокращении агонистов и растяжении антагонистов последние не сокращаются и становится возможно осуществление рефлекса.
По классической схеме после возбуждения рецептора сигнал центростремительно идёт по периферическому отростку к телу клетки спинномозгового ганглия, а затем проходит в составе задних корешков по центральному отростку. В результате получается смешанный нерв (как было описано ранее), чувствительные импульсы от которого направляются в задние рога спинного мозга, а проприоцептивные — в задние канатики, где отростки делятся на длинные и короткие ветви. Короткие волокна устанавливают межсегментарные связи и участвуют в реализации проприоцептивных рефлексов (проникают в серое вещество спинного мозга и достигают клеток переднего рога).
Источники:
Михайленко “Клиническая неврология: семиотика и топическая диагностика”, 2012 г.
Синельников “Атлас анатомии человека”, 4 том, 2015 г.
