
Дефицит адгезии лейкоцитов
При попадании микроорганизмов в ткань начинается воспалительный процесс. Одним из его следствий является элиминация лейкоцитами чужеродного патогена и поддержание таким образом тканевого гомеостаза. Чтобы это произошло, лейкоцитам нужно осуществить перемещения: сначала по сосудам, затем непосредственно в ткань, и лишь тогда расправиться с микроорганизмом. Осуществляется это с помощью разнообразных вспомогательных белковых молекул — т. н. медиаторов воспаления, а также с помощью различных адгезинов, необходимых для полноценного иммунного ответа. Врожденное отсутствие последних было названо «дефицитом адгезии лейкоцитов», и на сегодняшний день заболевание считается неизлечимым.
Дефицит адгезии лейкоцитов (ДАЛ) — редкое наследственное заболевание, крайне опасное из-за тяжелейшего врожденного иммунодефицита: фагоцитарные клетки неспособны проникнуть в ткани, оставляя таким образом клетки беззащитными перед вирусами и бактериями.
В настоящее время выделяют три формы ДАЛ: ДАЛ-1, ДАЛ-2 и ДАЛ-3 (последнюю также называют «вариантом ДАЛ-1»). Чтобы разобраться в этом, сначала следует проследить механизм перехода лейкоцитов в ткань.
Механизм проникновения лейкоцитов в очаг воспаления
Лейкоциты активируются в фазу альтерации различными хемоаттрактантами (из группы медиаторов воспаления), которые в дальнейшем способствуют миграции лейкоцитов и их последующему выходу в ткани. Выделяют две группы аттрактантов:
— «Классические» аттрактанты — N-формильные бактериальные пептидные антигены [1], белки системы комплемента С3а и С5а, а также различные липидные молекулы по типу лейкотриена В4 [2].
— Т. н. «селективные» аттрактанты — группа белков суперсемейства хемокинов, которые можно условно разделить на 4 группы: CXC, CX3C, CC и C (названы так по наличию двух первых остатков цистеина — парных и высококонсервативных) [2], [3].
Хемоаттрактанты связываются со специфическими рецепторами лейкоцитов и вызывают их перемещение по «градиенту хемоаттракции», то есть к месту очага воспаления. У человека известно около 18 хемокиновых рецепторов, все они связаны с G-белками и состоят из 7 трансмембранных доменов, что является типичным для многих рецепторов [2]. При активации этих рецепторов начинает синтезироваться группа молекул адгезии, которая необходима для последовательного каскадного перехода лейкоцитов в ткань.
Изначально лейкоциты движутся с током крови, постепенно выходя из осевого потока на периферию и приближаясь к стенке сосуда (маргинация), после чего лейкоциты прикрепляются к эндотелию и просачиваются сквозь него непосредственно к месту повреждения.
Классически выделяют три этапа в процессе достижения тканевого очага воспаления [4]: «качание», обусловленное селектинами; активация лейкоцитов под влиянием хемокинов и внедрение в ткань, опосредованное интегринами.
В настоящее время эти этапы несколько расширены (см. рис.1) [5]. Первым этапом остается «качание» или «перекатывание» лейкоцитов, обусловленное особыми белками — селектинами и интегинами.
Селектины можно назвать молекулами распознавания и прикрепления. Большинство лейкоцитов секретируют селектин P, клетки эндотелия (воспаленные) — селектины Е и L. Основной рецептор для всех них — P-селектиновый гликозилированный рецептор-1 (PSGR-1). Именно за счет него лейкоциты способны прочно прикрепиться к эндотелию; благодаря ему же лейкоциты, которые не обладают необходимым «рецепторным паспортом», не могут пройти в очаг воспаления.
Интегрины — еще одна группа белков, обладающих адгезивными свойствами. В их группе выделяют два основных подсемейства: β1- и β2-интегрины, также имеются и другие. Наиболее важными являются следующие представители:
- α4β7-интегрин — необходим для прикрепления к рецепторам клеточной адгезии слизистой сосуда MADCAM-1 (англ. mucosal vascular addressin cell-adhesion molecule-1);
- α4β1-интегин или VLA4 (англ. very late antigen-4) — необходим для прикрепления к внеклеточному рецептору адгезии VCAM-1 (англ. vascular cell-adhesion molecule-1);
- β2-интегрин — необходим для соединения с внутриклеточным рецептором адгезии ICAM-1 (англ. intracellular adhesion molecule-1).
- Также имеется αLβ2-интегрин, имеющий аббревиатуру LFA-1( lymphocyte function-associated antigen-1), который также связывается с ICAM-1 рецептором [5].
Все эти сложности нужны лейкоциту для того, чтобы перейти в фазу «медленного качания», а затем и вовсе остановиться. «Захват» лейкоцитов происходит путем прочного связывания интегринов с белками VCAM и ICAM (их относят к суперсемейству иммуноглобулинов, синтезируются эндотелиоцитами под влиянием медиаторов воспаления) [6].
Чтобы произошло прочное связывание (адгезия), также нужны молекулы-регуляторы — уже знакомые нам хемокины (основные — ИЛ-4, ФНО). Они могут содержаться в кровотоке, транспортироваться эндотелиоцитами или синтезироваться активированными тромбоцитами и тучными клетками. Также эти хемокины обладают высокой аффинностью к упомянутым рецепторам, ассоциированным с G-белком (GPCR – G-protein coupled receptor) [5].
Условно считается, что β1-интегрины — молекулы эозинофилов, моноцитов и лимфоцитов, связываются с VCAM, а β2- интегрины — белки нейтрофилов и базофилов – с ICAM [22]; с их помощью лейкоцит прочно прикрепляется к эндотелию. После этого начинается финальный этап — трансмиграция.
Проникновение через стенку сосуда — чрезвычайно сложный процесс. Начинается он с медленного перемещения или «ползания» фагоцита по эндотелию (с помощью макрофагального антигена 1 — MAC1, рис.1). Лейкоцит «ищет» удобное место для миграции в ткань. Мигрировать лейкоцит способен как через межклеточное пространство (в большинстве случаев), так и непосредственно через клетку (таков механизм трансмиграции лейкоцитов в ЦНС) [5].
Между эндотелиоцитами лейкоцит проникает благодаря крайне гибкой мембране, он буквально «просачивается» сквозь межклеточные соединения. Процесс этот довольно длителен и сложен, занимает 15–20 минут и регулируется множеством хемокинов, цитокинов и др. [5, 22].
Трансцеллюлярно (непосредственно через эндотелиоцит) фагоцит проходит с помощью недавно открытого транспортного механизма — везикуло-вакуолярных органелл (VVO) [7]. Это небольшие мембранные структуры, предположительно специализированные транспортеры лейкоцитов (а именно нейтрофилов) через тело эндотелиоцита.
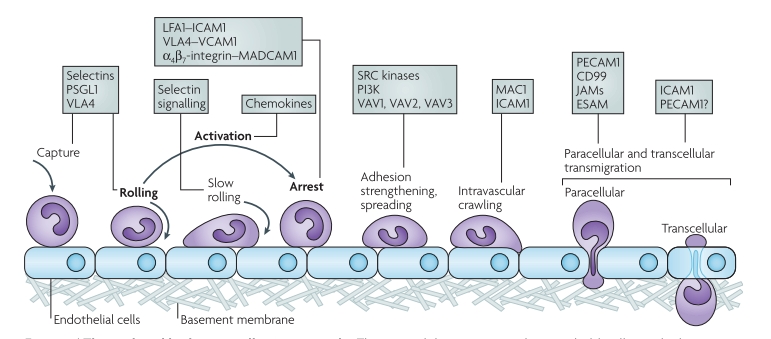
Рисунок 1 | Поэтапное проникновение лейкоцита через эндотелий сосуда [5].
Наконец, преодолев эндотелий, фагоцитарная клетка оказывается в очаге воспаления, где наводит порядок и способствует восстановлению ткани.
Из описанного выше механизма активации лейкоцитов и проникновения их в очаг воспаления становится понятна важность правильной работы всех задействованных молекул. Но вследствие определенной генетической мутации адгезивная способность лейкоцитов может быть нарушена — такое состояние называется дефицитом адгезии лейкоцитов (ДАЛ).
Классифицируют ДАЛ исключительно по виду дефектных молекул (и генов, их кодирующих). Клиническая картина, подход к диагностике и возможному лечению (или купированию заболевания) одинаковы в общих чертах, хотя имеются и нюансы.
Дефицит адгезии лейкоцитов 1 типа
ДАЛ 1 типа — редкое заболевание, передающееся по аутосомно-доминатному типу наследования. Этот первичный иммунодефицит кодируется геном ITGB2, отвечающим за экспрессию β2-интегринов (CD11/CD18). Распространенность заболевания — 1:1 000 000, в литературе можно встретить всего около 300 случаев [8], [9].
Впервые о нем сообщили как о полноценном самостоятельном заболевании в 1997 году [10], хотя в литературе и ранее были описаны случаи иммунодефицитов по причине дефекта β2-интегринов. Больше всего страдают нейтрофилы, которые не способны проникнуть в ткань.
Клинически это проявляется рецидивирующими жизнеугрожающими бактериальными и грибковыми инфекциями [11], [12]. Зачастую иммунодефицит проявляет себя очень рано, причем инфекции быстро переходят в септическую форму.
Отличительная особенность любого ДАЛ — отсутствие гноя в очаге воспаления. На коже и слизистых обнаруживаются очаги некроза, в детском возрасте смертность крайне высока. В дальнейшем (если грамотно устранять все инфекции) болезнь проявляет себя не так остро, более взрослые пациенты страдают от тяжелых гингивитов и хронических периодонтитов, но вероятность летального исхода у них существенно ниже.
Диагностика может осуществляться методами генной инженерии, но для ДАЛ-1 это нецелесообразно. Гораздо чаще выявляют маркеры CD11/CD18 методом проточной цитометрии — привычный и даже рутинный анализ в иммунологии [8]. Соответственно, при отсутствии данных маркеров можно говорить о ДАЛ 1 типа.
Единственным методом лечения ДАЛ-1 на данный момент является аллогенная трансплантация гемопоэтических клеток (АТГК) [13]. На протяжении с 1993 по 2007 год АТГК была произведена 36 пациентам в возрасте от 2 мес до 14 лет (средний возраст — 9 месяцев). Выживаемость составила около 75 %. Важно отметить, что большая часть смертей произошла по причине невозможности найти HLA-совместимого донора.
Помимо этого, существует достаточно перспективная патогенетическая терапия, а именно — трансфузия гранулоцитов. Метод на сегодняшний день достаточно спорный, разумеется, устранить ДАЛ он неспособен, зато вполне может справиться с тяжелыми инфекциями, как, например, с гангренозной эктимой [14].
Наконец, ДАЛ-1 можно купировать и сдерживать с помощью грамотной антибиотикотерапии, причем это тот самый случай, когда антибиотики следует применять и в качестве профилактики [11].
Дефицит адгезии лейкоцитов 2 типа
В основе ДАЛ-2 — дефект селектинов и, как следствие, невозможность «перекатывания» лейкоцитов по эндотелию. Но, в отличие от ДАЛ-1, этиология иммунодефицита не так очевидна, хотя бы потому, что известно всего лишь около 10 случаев данного заболевания [11].
Молекулярная природа ДАЛ-2 — недостаточность белка-переносчика ГДФ-фукозы (фукозилирование — одна из необходимых модификаций трансмембранных рецепторов адгезии [15]); сам белок переносчик — GFTP (англ. GDP-fucose transport protein) кодируется геном SLC35C1, он же FUCT1 [16].
Также дефицит вышеупомянутого GFTP приводит к еще двум редким феноменам: нарушение синтеза антигенов ABO-системы (бомбейский феномен) и недостаточность антигенов системы Льюиса [17]. Таким образом, дефект одного гена FUCT1 приводит сразу к трем редчайшим патологиям, которые крайне тяжело поддаются корректировке.
Клиническая картина, как ни странно, зачастую более мягкая в сравнении с ДАЛ1. Лейкоциты, хотя и лишены правильно работающих селектинов, все же имеют абсолютно интактные интегрины, то есть, хотя бы какая-то часть фагоцитов все же способна проникает в ткань и оказывает защиту от инфекции, которые становятся менее серьезными, чем в предыдущем случае. Иммунодефицит проявляется в основном на первом году жизни, в более взрослом периоде осложнения не так выражены (хотя и остается хронический периодонтит) [11].
Однако же не все так хорошо, как хотелось бы. Недостаточность все того же транспортера GFTP может проявляться (хоть и редко) задержкой роста и умственной отсталостью [18].
Диагностировать ДАЛ-2 можно на основании сочетанного феномена бомбейской группы крови и отсутствия антигенов Льюиса; окончательным же доказательством существования патологии будет генетический анализ гена FUCT1 [11].
Лечение ДАЛ-2 обычно протекает безрезультатно. На сегодняшний день помимо симптоматического лечения антибиотиками медицине предложить, в общем-то, нечего. Единственным более-менее эффективным может быть прием L-фукозы орально. По крайней мере, в исследовании состояние пациентов кратковременно улучшилось; однако спустя месяц нейтрофилы перестали реагировать на подобную поддержку [19].
Дефицит адгезии лейкоцитов 3 типа
Часто ДАЛ-3 относят к разновидностям ДАЛ-1 типа, однако такой подход вряд ли правильный. ДАЛ-3 гораздо более агрессивное заболевание, которое практически невозможно купировать.
Этиология болезни заключается в дефекте гена FERMT3, который отвечает за синтез всех β-интегринов. Результатом становится полная неспособность лейкоцита к хоть какому-нибудь прикреплению к эндотелиоцитам, не говоря уже о проникновении через стенку сосуда [11].
Помимо обычных тяжелых инфекций больные страдают также геморрагическим синдромом (наподобие тромбастении Гланцмана — продолжительность кровотечения составляет более 15 минут при норме 2–6 мин [20]. Это вполне ожидаемо, так как тромбоциты в процессе агрегации используют β3-интегрины.
ДАЛ-3 на сегодня выявлен примерно у 20 семей во всем мире. Это крайне тяжелая форма ДАЛ с высокой летальностью, зачастую новорожденные в таких семьях очень быстро умирали, причем еще до постановки диагноза.
В дополнение к уже описанной клинической картине (рецидивирующие тяжелые инфекции, отсутствие гноя, геморрагический синдром) можно назвать дефект костной ткани наподобие остеопетроза (остеосклероз, мраморная болезнь) [21]. Выявляется не у всех пациентов, однако подобная патология может существенно помочь в постановке диагноза.
Собственно диагностика проводится с помощью генного анализа. Существует ряд тестов, в которых можно распознать дефект нейтрофилов, например, тест с неопсонизированным зимозаном [11], который поможет дифференцировать ДАЛ-3 от ДАЛ-1. Однако подобные тесты требуют времени, а заболевание зачастую не оставляет шанса на раздумья.
Лечение мало отличается от такового при других формах ДАЛ. Это трансфузии гранулоцитов и (в данном случае) тромбоцитов, причем число трансфузий может достигать 50 в год. Единственным же способом этиотропной терапии остается аллогенная трансплантация гемопоэтических клеток [11].
Источники:
- E. Schiffmann, B. A. Corcoran, and S. M. Wahl, “N-formylmethionyl peptides as chemoattractants for leucocytes.,” Proc. Natl. Acad. Sci., vol. 72, no. 3, pp. 1059–1062, 1975.
- C. Murdoch and A. Finn, “Chemokine receptors and their role in inflammation and infectious diseases,” Blood, vol. 95, no. 10, pp. 3032–3043, 2000.
- M. Baggiolini, B. Dewald, and B. Moser, “Human Chemokines: An Update,” Annu. Rev. Immunol., vol. 15, no. 1, pp. 675–705, 1997.
- E. C. Butcher, “Leukocyte-endothelial cell recognition: Three (or more) steps to specificity and diversity,” Cell, vol. 67, no. 6, pp. 1033–1036, 1991.
- K. Ley, C. Laudanna, M. I. Cybulsky, and S. Nourshargh, “Getting to the site of inflammation: The leukocyte adhesion cascade updated,” Nat. Rev. Immunol., vol. 7, no. 9, pp. 678–689, 2007.
- J. J. Campbell, J. Hedrick, A. Zlotnik, M. A. Siani, D. A. Thompson, and E. C. Butcher, “Chemokines and the arrest of lymphocytes rolling under flow conditions,” Science (80-. )., vol. 279, no. 5349, pp. 381–384, 1998.
- A. M. Dvorak and D. Feng, “The vesiculo-vacuolar organelle (VVO): A new endothelial cell permeability organelle,” J. Histochem. Cytochem., vol. 49, no. 4, pp. 419–431, 2001.
- Hamid Nawaz Tipu, “Leukocyte Adhesion Deficiency Type I: A Rare Primary Immunodeficiency Disorder,” Pediatr. Allergy Immunol., vol. 28, no. 3, pp. 303–305, 2017.
- E. Almarza Novoa et al., “Leukocyte adhesion deficiency-I: A comprehensive review of all published cases,” J. Allergy Clin. Immunol. Pract., vol. 6, no. 4, p. 1418–1420.e10, 2018.
- T. W. Kuijpers et al., “Leukocyte Adhesion Deficiency Type 1/Variant: Dysfunctional b 2 Integrins,” J. Clin. Invest., vol. 100, pp. 1725–1733, 1997.
- E. van de Vijver, T. K. van den Berg, and T. W. Kuijpers, “Leukocyte Adhesion Deficiencies,” Hematol Oncol Clin N Am., vol. 27 (1), pp. 101–116, 2013.
- M. Movahedi et al., “Clinical and laboratory findings in Iranian patients with leukocyte adhesion deficiency (study of 15 cases),” J. Clin. Immunol., vol. 27, no. 3, pp. 302–307, 2007.
- W. Qasim et al., “Allogeneic Hematopoietic Stem-Cell Transplantation for Leukocyte Adhesion Deficiency,” Pediatrics, vol. 123, no. 3, pp. 836–840, 2009.
- F. Mellouli et al., “Successful treatment of fusarium solani ecthyma gangrenosum in a patient affected by leukocyte adhesion deficiency type 1 with granulocytes transfusions,” BMC Dermatol., vol. 10, pp. 2–4, 2010.
- B. Ma, J. L. Simala-Grant, and D. E. Taylor, “Fucosylation in prokaryotes and eukaryotes,” Glycobiology, vol. 16, no. 12, 2006.
- K. Lühn, M. K. Wild, M. Eckhardt, R. Gerardy-Schahn, and D. Vestweber, “The gene defective in leukocyte adhesion deficiency II encodes a putative GDP-fucose transporter,” Nat. Genet., vol. 28, no. 1, pp. 69–72, 2001.
- A. Hidalgo, S. Ma, A. J. Peired, L. A. Weiss, C. Cunningham-Rundles, and P. S. Frenette, “Insights into leukocyte adhesion deficiency type 2 from a novel mutation in the GDP-fucose transporter gene,” Blood, vol. 101, no. 5, pp. 1705–1712, 2003.
- T. Marquardt et al., “Leukocyte adhesion deficiency II syndrome, a generalized defect in fucose metabolism,” J. Pediatr., vol. 134, no. 6, pp. 681–688, 1999.
- S. Yakubenia and M. K. Wild, “Leukocyte adhesion deficiency II: Advances and open questions,” FEBS J., vol. 273, no. 19, pp. 4390–4398, 2006.
- K. Jurk et al., “Novel integrin-dependent platelet malfunction in siblings with leukocyte adhesion deficiency-III (LAD-III) caused by a point mutation in FERMT3,” Thromb. Haemost., vol. 103, no. 5, pp. 1053–1064, 2010.
- S. Schmidt et al., “Kindlin-3-mediated signaling from multiple integrin classes is required for osteoclast-mediated bone resorption,” J. Cell Biol., vol. 192, no. 5, pp. 883–897, 2011.
- E.F. Goljan, “Pathology”, Chapter 3, 2012.
