
Эпигенетические механизмы наркозависимости

В настоящее время существует мало фармакологических методов лечения наркомании, а те, что есть, имеют низкую эффективность у многих людей. Это может быть связано с постоянными функциональными изменениями в головном мозге, индуцированными приемом наркотиков и лекарств, индивидуальным фенотипом зависимости. Постоянный прием наркотиков, связанный с зависимостью, вызывает изменения в структуре и функции клеток и путей головного мозга, которые лежат в основе аддиктивного поведения, например, поиск наркотиков и склонность к рецидивам. Поэтому определение целевых механизмов, которые управляют функциональными изменениями работы мозга, это важный шаг в изучении этиологии зависимости и разработки новых методов лечения. Это потребует всестороннего понимания нейробиологических процессов зависимости, включая роль экспрессии генов и регуляцию экспрессии, изменения в структуре и функции нейронов, индуцированные приемом наркотиков [1].
Термин «эпигенетика» используется для описания динамических молекулярных модификаций, отражающихся на хроматине в ядре клетки, что регулирует функции таких процессов, как репарация ДНК, организация хроматина, транскрипция и сплайсинг РНК и другие. Ученые, исследующие явление наркомании, заинтересовались изучением эпигенетики из-за того факта, что индивидуальный опыт, в частности волевое, многократное употребление наркотиков, изменяет конфигурацию хроматина в мозге, что вариативно в зависимости от области мозга и типа клетки [2, 6].
Эпигенетические механизмы действуют на протяжении всей жизни. Такие механизмы, вероятно, важны на ранних этапах развития, так как вносят свой вклад в различия между индивидуумами, которые возникают случайно даже при одинаковых условиях среды. Когда определенные типы эпигенетических модификаций после их возникновения являются постоянными, возможно, что определенные эпигенетические изменения, возникающие в результате поведенческого опыта или случайных событий развития, лежат в основе постоянных изменений в функции мозга [3].
Нейроэпигенетический ландшафт
Считается, что путем регулирования процессов, связанных с ДНК, эпигенетические изменения, индуцированные веществами, способствуют нарушению клеточных функций, чем и объясняется патогенез наркомании. Есть многообещающий терапевтический потенциал в отношении нацеливания на ключевые эпигенетические модификации для борьбы с привыканием [2].
Посттрансляционные модификации (ПТМ) гистонов меняют пространственную структуру хроматина, контролируя процессы, связанные с ДНК. Гистоновые субъединицы могут быть модифицированы путем ацетилирования, метилирования, фосфорилирования, рибозилирования АДФ, убиквитилирования и сумоилирования и др. Гистоновые ПТМ обратимы: они динамически осуществляются записывающими белками, которые распознаются считывающими белками, которые опосредуют клеточный ответ, и удаляются стирающими белками. Экспрессия и функция многочисленных записывающих белков, стирающих и считывающих изменяются как у людей с зависимостью, так и в животных моделях зависимости. Восстановление нормальной функции этих белков с помощью фармакотерапии — новая ниша для разработки лекарств против наркомании [2].
.
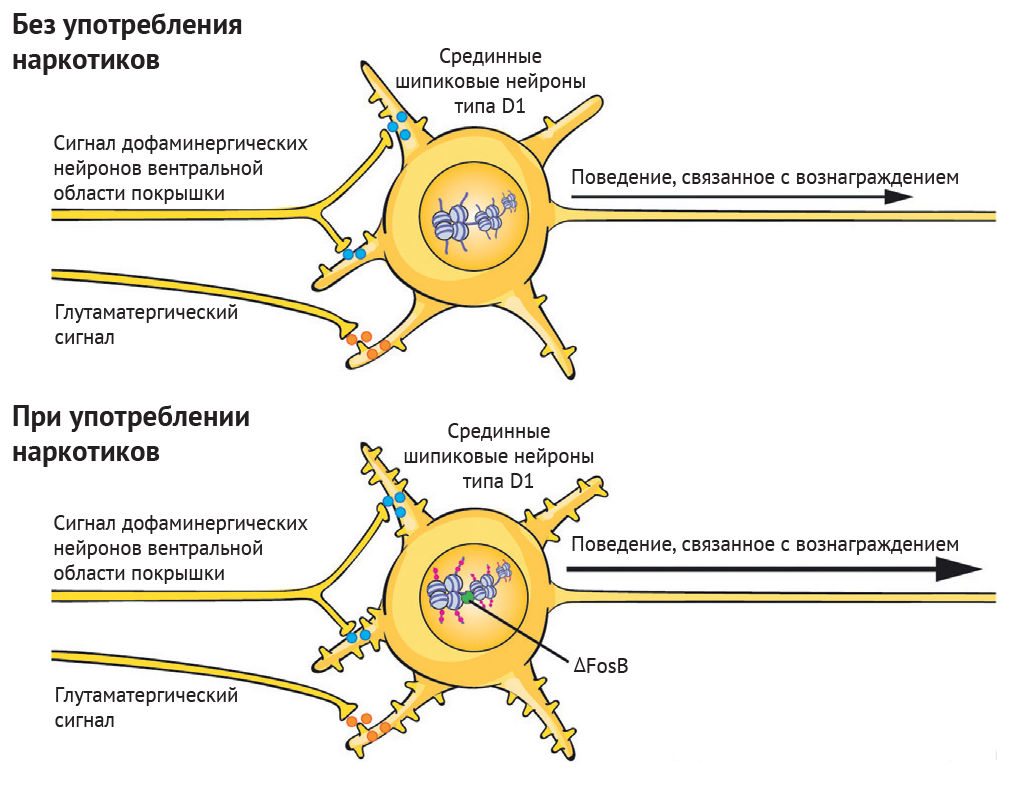
Внизу: хроническое употребление наркотиков нарушает баланс регулирующих белков, записывающих и стирающих, что приводит к эпигенетическим адаптациям в определенных локусах в ядре срединных шипиковых нейронов.
Адаптации и лекарственная индукция факторов транскрипции (например, DFosB) опосредуют изменения транскрипции многих генов, включая гены, кодирующие нейротрансмиттерные рецепторы, белки цитоскелета и ионные каналы. Вследствие этих транскрипционных адаптаций меняется морфология срединных шипиковых нейронов (например, показана повышенная плотность дендритных шипиков) и физиологическая функция в отношении процессов вознаграждения. Это лежит в основе поведенческих дезадаптаций, определяющих зависимость.
Синаптическая пластичность в контексте наркозависимости
Синаптическая пластичность — это возможность изменения силы синапса (величины изменения трансмембранного потенциала) в ответ на активацию постсинаптических рецепторов [4].
Начальная доза наркотического препарата потенцирует возбуждающие афферентные волокна на дофаминовые нейроны вентральной области покрышки. Потенцирование возбуждающих глутаматергических афферентов из медиальной префронтальной коры и вентрального гиппокампа на срединные шипиковые нейроны прилежащего ядра, экспрессирующие рецептор D1, ассоциировано с поиском наркотиков. Для индукции такой пластичности обычно требуется дофамин, механизмы экспрессии варьируются, и метаботропные рецепторы глутамата могут ограничивать потенцирование. Для возбуждающей передачи характерной особенностью является инсерция глутаматных рецепторов AMPA — и в некоторых случаях инсерция проницаемых для кальция AMPA-рецепторов без GluA2 в постсинаптическую плазматическую мембрану. Вызванная лекарственным средством пластичность передачи ГАМК выражается пресинаптическим механизмом, изменяющим высвобождение ГАМК. Нейроны прилежащего ядра также экспрессируют проницаемые для кальция AMPA-рецепторы после воздействия лекарственного средства, особенно при употреблении кокаина [3,5].
.
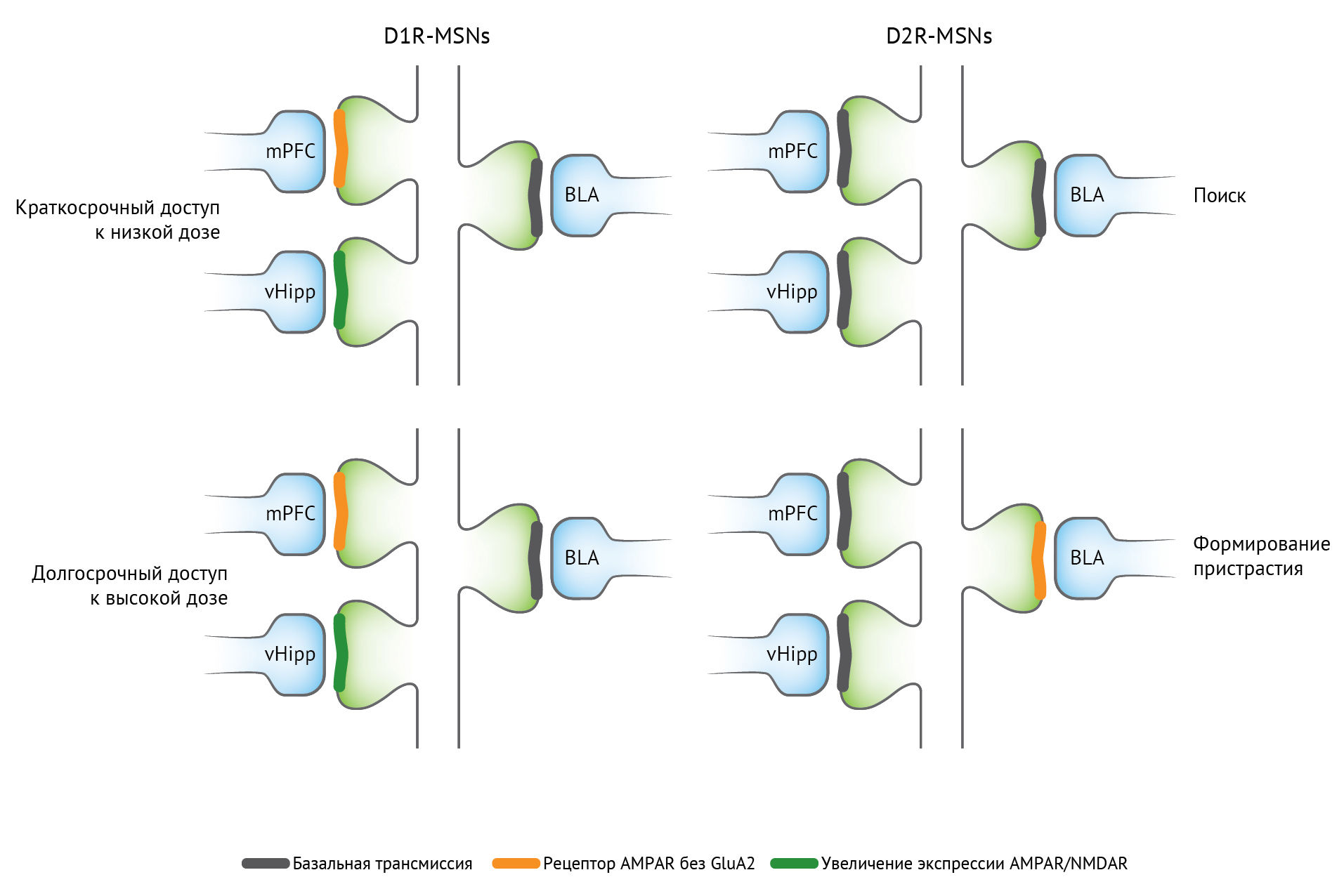
Сокращения: AMPAR — рецептор α-амино-3-гидрокси-5-метил-4-изоксазолпропионовой кислоты; БЛА — базолатеральная миндалина; D1R — дофаминовый рецептор — D1 рецептор; D2R — дофаминовый рецептор D2; mPFC — медиальная префронтальная кора; MSN — срединный шипиковый нейрон; NMDAR — N-метил-D-аспартатный рецептор; vHipp — вентральный гиппокамп.
Воздействие кокаина и опиатов регулирует общее количество функциональных глутаматергических синапсов на прилежащем ядре срединных шипиковых нейронов, так как молчащие синапсы экспрессируют NMDA-рецептор и не экспрессируют AMPA-рецептор [3].
АМРА-рецептор (рецептор α-амино-3-гидрокси-5-метил-4-изоксазолпропионовой кислоты, AMPAR), релокализованные после первого воздействия наркотических средств, заменяются на GluA2-содержащие рецепторы, которые синтезируются de novo. В прилежащем ядре сопутствующая активация рецепторов D1R и N-метил-D-аспартата (NMDAR) запускает путь MAP-киназы-ERK, который влияет на транскрипцию. Ремоделирование путей прилежащего ядра, лежащее в основе привычек и зависимостей, и нескольких областей выше, иннервирующих прилежащее ядро через глутаматергические нейроны — префронтальная кора, вентральный гиппокамп, базолатеральная миндалина и таламус, каждый из которых получает дофамин из дофаминовых нейронов вентральной области покрышки — появляются в качестве основных локусов ремоделирования путей дофамина. Область, которой уделяется наибольшее внимание — медиальная префронтальная кора, с нисходящими глутаматергическими путями от медиальной префронтальной коры до прилежащего ядра и нескольких других подкорковых областей, связанных с неадаптивным поведением и индивидуальной уязвимостью (рисунок 3) [3].
.
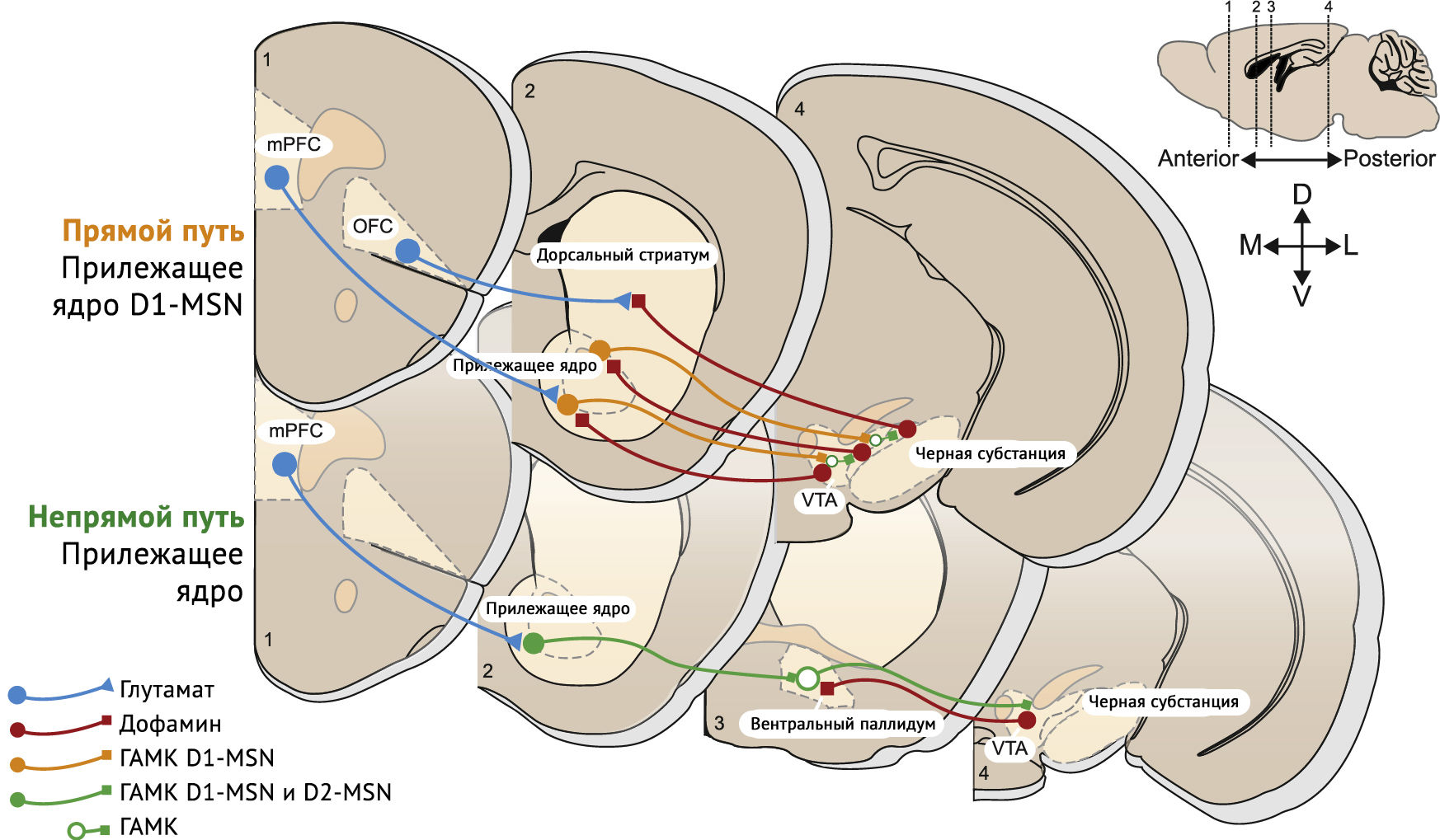
Синаптическое потенцирование наблюдалось в афферентных волокнах проекционных срединных шипиковых нейронов D1 и D2 и опосредовано постсинаптическим механизмом экспрессии. Механизмы индукции этих синаптических изменений недостаточно изучены. При хроническом воздействии наркотиков может быть задействовано все больше и больше нервных узлов и путей. Фактически анатомические знания и эксперименты подтверждают эту концепцию [3].
Стимуляция более дорсальной предлимбической области способствует употреблению наркотиков, тогда как стимуляция более вентральной инфралимбической области сдерживает рецидив после вымирания. Обе области могут вести и сдерживать поиск наркотиков в зависимости от текущей ситуации и исходных данных пациента. Усовершенствованная модель учитывает проекции путей отдельных нейронов медиальной префронтальной коры/прилежащего ядра, которые объединяются в прелимбической области и инфралимбической, чтобы достичь ядра и оболочки прилежащего ядра. При повторении приема нракотиков активность инфралимбической превалирует над активностью прелимбической области, а инактивация инфралимбической области восстанавливает целенаправленное поведение. Эта модель предполагает, что привычные показатели достигнуты при переключении с прелимбической области на инфралимбическую. Были вовлечены и другие области префронтальной коры — дисфункция орбитофронтальной коры, которая может способствовать неконтролируемому потреблению наркотических веществ. Если медиальная префронтальная кора и орбитофронтальная кора играют роль в обновлении аффективной ценности стимулов и результата действия во время целенаправленного поведения, они могут дисфункции в патологических состояниях, где зависимость является ключевым симптомом [3].
Развитие наркозависимости начинается с первого приема и постепенно консолидируется во время повторного, но все еще контролируемого употребления наркотиков. По мере того как потребление растет, прием наркотиков становится необходимым, что приводит к потере контроля над употреблением наркотиков. Это развитие может зависеть от формирования привычки, которое постепенно становится все более и более выраженным, что в конечном итоге квалифицируется как зависимость [3].
Источники:
1. Eagle A., Al Masraf B., Robison A. J. Transcriptional and Epigenetic Regulation of Reward Circuitry in Drug Addiction //Neural Mechanisms of Addiction. – Academic Press, 2019. – С. 23-34.
2. Hamilton P. J., Nestler E. J. Epigenetics and addiction //Current opinion in neurobiology. – 2019. – Т. 59. – С. 128-136
3. Nestler E. J., Lüscher C. The molecular basis of drug addiction: linking epigenetic to synaptic and circuit mechanisms //Neuron. – 2019. – Т. 102. – №. 1. – С. 48-59.
4. Citri A., Malenka R. C. Synaptic plasticity: multiple forms, functions, and mechanisms //Neuropsychopharmacology. – 2008. – Т. 33. – №. 1. – С. 18.
5. Lüscher C. The emergence of a circuit model for addiction //Annual review of neuroscience. – 2016. – Т. 39. – С. 257-276.
6. Bastle R. M., Neisewander J. L. Epigenetics and Drug Abuse //Recent Advances in Drug Addiction Research and Clinical Applications. – 2016. – С. 21
