
Троица COVID-19: иммунитет, воспаление и вторжение

Аннотация
Коронавирус-2 (SARS-CoV-2), сопряженный с тяжелым острым респираторным синдромом, является возбудителем текущей пандемии коронавирусной болезни 2019 года (COVID-19). Наряду с исследованиями вирусологии SARS-CoV-2, понимание фундаментальных физиологических и иммунологических процессов, лежащих в основе клинических проявлений COVID-19, жизненно важно для идентификации вируса и рационального дизайна эффективных методов лечения. В данном обзоре будет рассмотрена патофизиология инфекции SARS-CoV-2. Мы описываем взаимодействие SARS-CoV-2 с иммунной системой и последующий вклад дисфункциональных иммунных реакций в прогрессирование заболевания. Помимо анализа сообщений, описывающих течение SARS-CoV-2, мы делаем выводы на основе патофизиологических и иммунологических особенностей других коронавирусов человека, преимущественно поражающих нижние дыхательные пути, вызывающих тяжелый острый респираторный синдром (SARS-CoV) и ближневосточный респираторный синдром (MERS-COV).
Первые случаи коронавирусной болезни 2019 (COVID-19), вероятно, произошли в Китае в декабре 2019 года в результате зоонозной передачи, связанной с крупным рынком морепродуктов, на котором также торговали живыми дикими животными. Возбудитель — коронавирус-2, сопряженный с острым респираторным синдромом (SARS-CoV-2), — передается от человека человеку, в связи с чем он быстро распространился в другие части Китая, а затем и на другие континенты. На 24 марта 2020 года SARS-CoV-2 заразил более 381 000 человек в 195 странах / регионах и убил более 16 000 (на 22 мая 2020 года насчитывается 5 125 612 зараженных и 333 382 летальных исходов — прим. ред.), в результате чего Всемирной организацией здравоохранения была объявлена пандемия [1]. Ежедневные сообщения о резком росте числа новых случаев по-прежнему поступают из многих стран и регионов, но меры по преодолению вируса не столь эффективны из-за отсутствия знаний о некоторых важных аспектах инфекции SARS-CoV-2: от биологии патогенного микроорганизма до реакции организма хозяина и вариантов лечения.
Таким образом, существует неотложная необходимость в лучшем понимании биологии взаимодействия хозяин-патоген при COVID-19, поскольку это даст важную информацию для определения тактики ведения заболевших, включая установление новых методов лечения. Здесь мы рассмотрим литературу по патофизиологии SARS-CoV-2, его взаимодействию с клетками-мишенями и иммунному ответу на вирус, включая вклад дисфункционального иммунного ответа в прогрессирование заболевания. В частности, мы подчеркиваем значение специфических особенностей инфекции для разработки многообещающих методов лечения, которые могут быть направлены на вирус или дисфункциональный иммунный ответ. Кроме того, исследования, посвященные адаптивному иммунному ответу, будут иметь решающее значение для разработки вакцин и препаратов моноклональных антител.
Патогенез COVID-19
Известно, что коронавирусы вызывают заболевания у людей и животных. Четыре из них (человеческие коронавирусы 229E, NL63, OC43 и HKU1) обычно поражают только верхние дыхательные пути и вызывают относительно легкие симптомы [2]. Однако существует три коронавируса (коронавирус тяжелого острого респираторного синдрома (SARS-CoV), коронавирус ближневосточного респираторного синдрома (MERS-CoV) и SARS-CoV-2, которые могут реплицироваться в нижних дыхательных путях и вызывать пневмонию с вероятностью смертельного исхода. SARS-CoV-2 относится к роду бета-коронавирусов. Его ближайший родственник среди коронавирусов человека — SARS-CoV, с 79 % генетического сходства [3]. Однако среди всех известных генетических последовательностей коронавирусов SARS-CoV-2 наиболее похож на коронавирус летучей мыши RaTG13 со сходством 98 % [4]. Также высокое сходство наблюдается с геномом коронавируса панголина (чешуйчатый муравьед) [5].
Как и другие респираторные коронавирусы, SARS-CoV-2 передается в основном воздушно-капельным путем, также возможен, но не доказан фекально-оральный путь передачи. Средний инкубационный период инфекции составляет приблизительно 4–5 дней до появления симптомов [6–9], у 97,5 % пациентов симптомы развиваются в течение 11,5 дней [8]. На момент госпитализации у пациентов с COVID-19 обычно наблюдается лихорадка и сухой кашель; реже пациенты также испытывают затрудненное дыхание, боли в мышцах и/или суставах, головную боль, головокружение, диарею, тошноту и кровохарканье [6, 10–15]. В течение 5–6 дней после появления симптомов вирусная нагрузка SARS-CoV-2 достигает своего пика, что значительно раньше, чем у схожего SARS-CoV, при котором вирусная нагрузка достигает пика примерно через 10 дней после появления первых симптомов [16–19]. Тяжелые случаи COVID-19 прогрессируют до острого респираторного дистресс-синдрома (ОРДС) в среднем за 8–9 дней от начала заболевания [11, 20].
Патофизиология развития инфекции SARS-CoV-2 очень похожа на таковую при заражении SARS-CoV: агрессивная воспалительная реакция с вовлечением дыхательных путей [21]. Следовательно, тяжесть заболевания у пациентов обусловлена не только вирусной инфекцией, но и реакцией организма хозяина. Корреляция степени тяжести с возрастом также, в целом, соответствует эпидемиологии SARS-CoV и MERS-CoV [6, 11, 14].
ОРДС, наблюдаемый при тяжелом течении COVID-19, характеризуется затрудненным дыханием и низким уровнем кислорода в крови [22]. В результате у некоторых пациентов могут развиться вторичные бактериальные и грибковые инфекции [14]. ОРДС может привести к дыхательной недостаточности, которая является причиной смерти в 70 % случаев летальных исходов COVID-19 [22]. Кроме того, мощный выброс цитокинов иммунной системой в ответ на вирусную инфекцию и/или вторичные инфекции могут привести к цитокиновому шторму и симптомам сепсиса, который является причиной смерти в 28 % летальных случаев COVID-19 [22], вследствие неконтролируемого воспаления, приводящего к полиорганной недостаточности, особенно поражая сердечную, печеночную и почечную системы (рис. 1). Большинство пациентов с SARS-CoV инфекцией, у которых развивалась почечная недостаточность, в конечном итоге умирали [23].
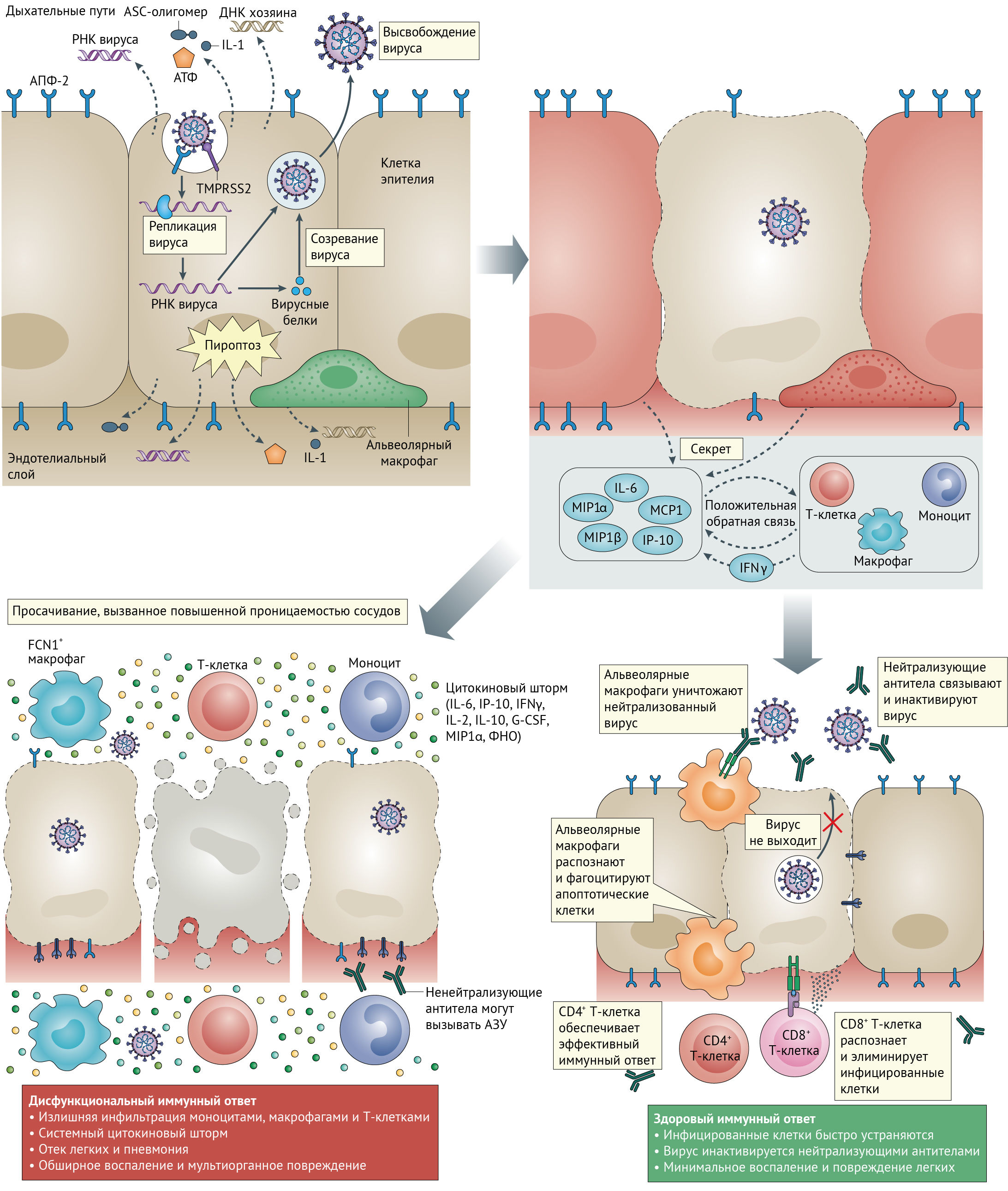
G-CSF — гранулоцитарный колониестимулирующий фактор;
ФНО — фактор некроза опухоли.
Инфицирование клетки-хозяина и его предупреждение
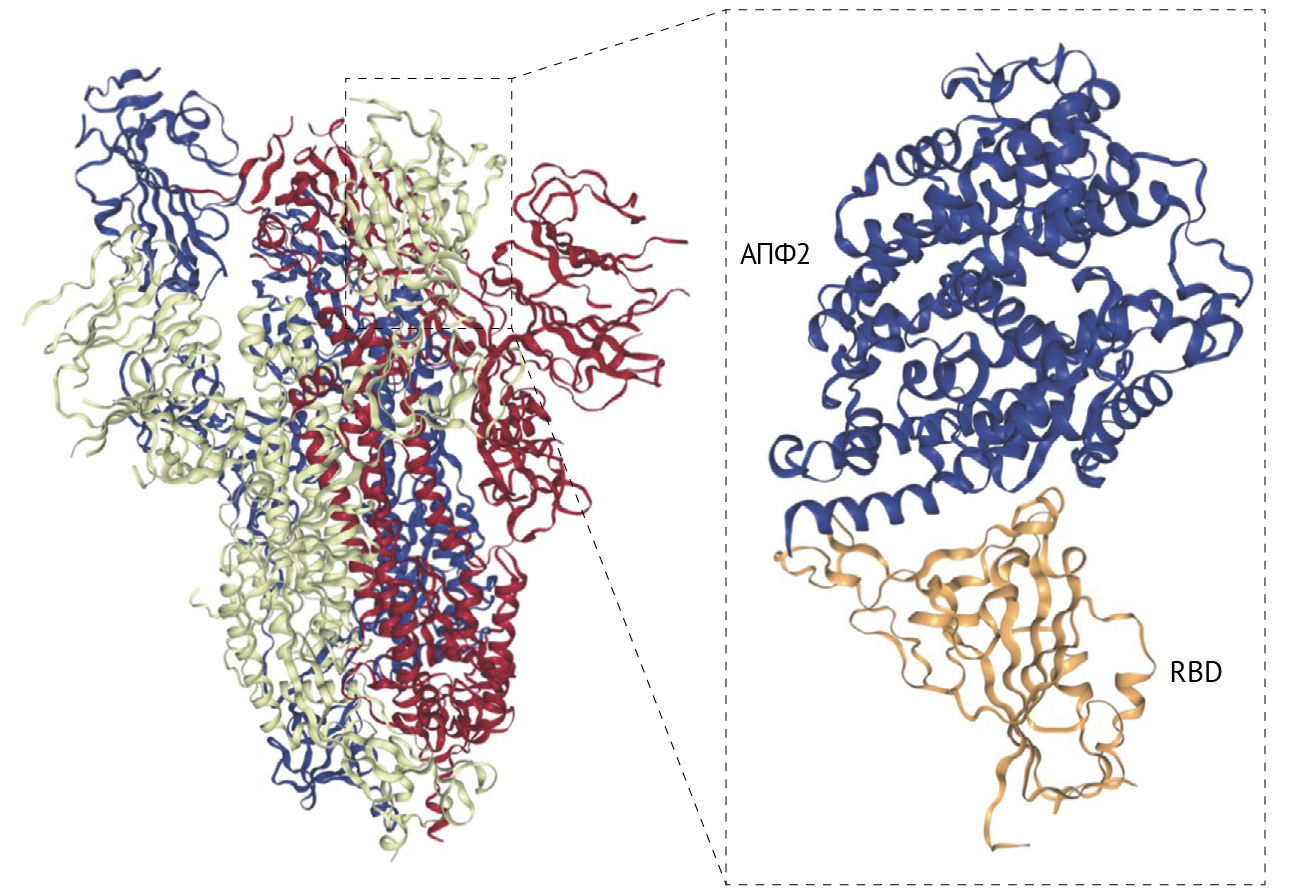
Первым этапом заражения является связывание вируса с клеткой-хозяином через рецептор-мишень. Ранее проведенная работа над исследованием SARS-CoV показала, что этот вирус главным образом нацелен на эпителиальные клетки дыхательных путей, альвеолярные эпителиальные клетки, эндотелиальные клетки сосудов и альвеолярные макрофаги, которые имеют на своей поверхности рецептор ангиотензин-превращающего фермента 2 (АПФ2), используемый SARS-CoV в качестве рецептора-мишени [24 –26] (рис. 2). Поскольку SARS-CoV-2 использует тот же рецептор проникновения, эти клетки, вероятно, являются также мишенью и для него [4, 27, 28]. Инфицирование SARS-CoV снижает экспрессию АПФ2 в клетках легких. Поскольку потеря функции АПФ2 в легких связана с острым повреждением легких, вызванное вирусом подавление АПФ2 может играть важную роль в патофизиологии заболевания [29–32]. Как было доказано, АПФ2 регулирует ренин-ангиотензиновую систему (РАС) [32]. Следовательно, снижение функции АПФ2 после инфицирования вирусом может привести к дисфункции РАС, оказывающей влияние на кровяное давление и водно-электролитный баланс, а также увеличивающей воспаление и проницаемость сосудов в дыхательных путях.
При COVID-19 наблюдается разница в уровне летальности между мужчинами (2,8 %) и женщинами (1,7 %) [33]. Поскольку АПФ2 расположен на Х-хромосоме, могут присутствовать аллели, которые придают устойчивость к COVID-19, что объясняет более низкую летальность среди женщин. Кроме того, половые гормоны эстроген и тестостерон имеют различные иммунорегуляторные функции, которые могут влиять на иммунную защиту и, следовательно, тяжесть заболевания [34].
SARS-CoV-2 имеет 79 % совпадения последовательности генома с SARS-CoV [4]. Шиповидный белок экспрессируется на поверхности вирусных частиц, придавая им характерный «коронованный» вид. Шиповидный белок состоит из двух субъединиц: S1 и S2. Субъединица S1 состоит из амино-терминального домена и рецептор-связывающего домена (RBD), который в SARS-CoV простирается от аминокислотного остатка 318 до аминокислотного остатка 510 [35–37]. RBD связывается с АПФ-2 клетки-хозяина как с рецептором-мишенью, запуская тем самым процесс инфицирования [4]. Связывание RBD с АПФ-2 запускает эндоцитоз вириона SARS-CoV-2 и подвергает его воздействию эндосомных протеаз [38]. Субъединица S2 состоит из области пептида слияния (FP) и двух областей гептадных повторов: HR1 и HR2 [39, 40]. Внутри эндосомы субъединица S1 отщепляется, обнажая пептид слияния, который внедряется в мембрану клетки-хозяина. Затем область S2 складывается сама для объединения областей HR1 и HR2. Это приводит к слиянию мембран и высвобождает вирусное содержимое в цитоплазму хозяина.
Существует 72 % сходства в аминокислотной последовательности RBD SARS-CoV и SARS-CoV-2, также у них очень похожи третичные структуры. Вычислительное моделирование и биофизические измерения показывают, что аффинность RBD SARS-CoV-2 к АПФ2 более высокая, чем у SARS-CoV [41, 42]. Кроме того, шиповидный белок SARS-CoV-2 содержит фурин-подобный сайт расщепления (аналогично MERS-CoV и человеческому коронавирусу OC43), который не обнаружен у SARS-CoV [43]. Эти характеристики могут способствовать повышенной контагиозности SARS-CoV-2 по сравнению с SARS-CoV. В дополнение к предварительному расщеплению фурином, для правильной обработки шиповидного белка SARS-CoV-2 и облегчения проникновения в клетку-хозяина также необходима клеточная сериновая протеаза TMPRSS2 [44].
Одним из путей разработки терапевтических средств против SARS-CoV-2 является блокирование целевого рецептора АПФ2 хозяина или TMPRSS2 (рис. 3). В настоящее время существуют соединения, нацеленные на эти молекулы, которые были одобрены для лечения других заболеваний. Например, алгоритмы нейросетей предсказывают, что барицитиниб, ингибитор янус-киназы (JAK), одобренный для лечения ревматоидного артрита, может ингибировать опосредованный АПФ2 эндоцитоз [45]. Другой ингибитор JAK, руксолитиниб, будет опробован в клинических испытаниях для лечения COVID-19 в конце этого года [46]. Альтернативная стратегия заключается в доставке высоких концентраций растворимой формы АПФ2, что потенциально может снизить проникновение вируса в клетки-мишени. Этот принцип тестируется с APN01, рекомбинантной формой АПФ2, разработанной APEIRON, которая в настоящее время проходит клинические испытания [47]. Моноклональные антитела к шиповидным белкам также могут ингибировать проникновение вируса в клетку и дополнительно обсуждаются в разделе «В-клеточный иммунитет».
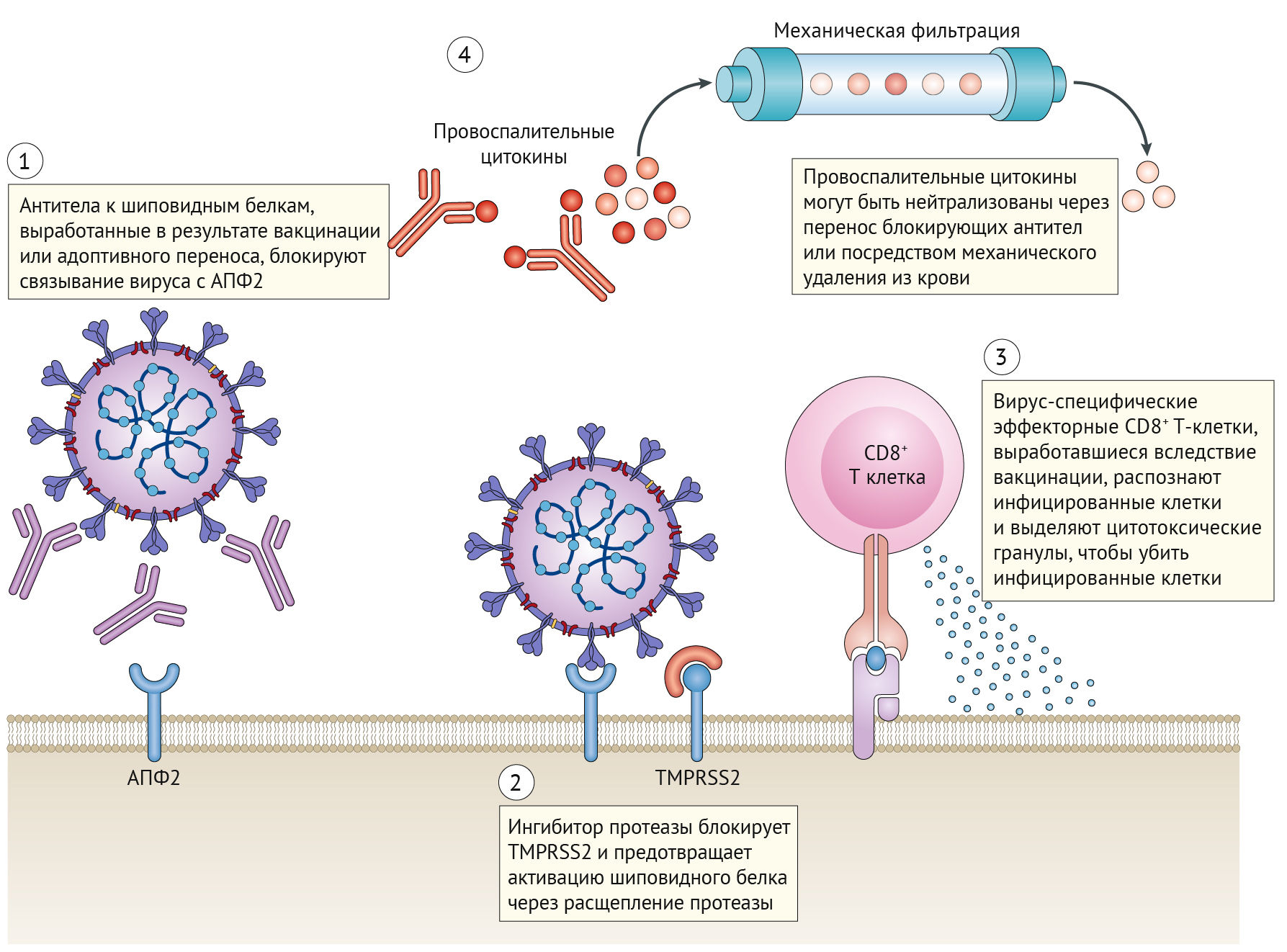
(2) Ингибиторы сериновой протеазы TMPRSS2 могут предотвращать расщепление шиповидного белка, которое является необходимым для внедрения вируса в клетку хозяина. Блокирование взаимодействия с АПФ2 или внедрения вируса также может предупредить инфицирование клетки-хозяина.
(3) Вирус-специфические CD8+ Т-клетки памяти, оставшиеся от предыдущей вакцинации или инфекции, могут дифференцироваться в эффекторные клетки во время повторного привлечения. Когда они идентифицируют инфицированные клетки, презентирующие вирус-специфические эпитопы, они дегранулируют и убивают инфицированные клетки, прежде чем те смогут производить зрелые вирионы.
(4) В новом методе лечения, который направлен на симптомы цитокинового шторма, кровь пациентов с коронавирусной болезнью 2019 года (COVID-19) может прогоняться через специализированные резервуары, которые специально разработаны для захвата провоспалительных цитокинов, после чего «очищенная» кровь возвращается пациентам.
Нафамостат мезилат [48, 49] и камостат мезилат [44] являются известными ингибиторами фермента TMPRSS2 и в настоящее время одобрены в нескольких странах для лечения других заболеваний. Хотя на момент написания статьи не проводилось никаких клинических испытаний, специально тестирующих эти препараты против COVID-19, когда мезилат камостата был протестирован на вирусе SARS-CoV-2, выделенном от пациента, он предотвращал проникновение вируса в клетки легкого [44, 50]. Если этот подход покажет свою эффективность, быстрое перепрофилирование таких препаратов будет рабочей и своевременной мерой в борьбе с COVID-19.
Воспалительный иммунопатогенез
Инфекция SARS-CoV-2 и разрушение клеток легких запускают местный иммунный ответ с привлечением макрофагов и моноцитов, которые отвечают на инфекцию, высвобождают цитокины и первичные адаптивные иммунные ответы T- и B-клеток. В большинстве случаев этот процесс способен привести к разрешению инфекции. Однако в некоторых случаях возникает дисфункциональный иммунный ответ, который может вызвать тяжелую легочную или даже системную патологию.
Цитопатические вирусы, включая SARS-CoV-2 [51], вызывают повреждение или гибель инфицированных вирусом клеток и тканей в ходе репликативного цикла вируса. Репликация вируса в эпителиальных клетках дыхательных путей [52] может вызывать высокий уровень связанного с вирусом пироптоза с ассоциированной повышеной проницаемостью сосудов, как это наблюдается у пациентов с SARS-CoV [53]. Пироптоз является воспалительной формой запрограммированной гибели клеток, которая обычно наблюдается при цитопатической вирусной инфекции [54]. Это вероятный триггер для последующего воспалительного ответа [55]. Во время инфекции SARS-CoV-2 повышается уровень IL-1β, важного цитокина, высвобождаемого во время пироптоза [11]. Используя различные паттерн-распознающие рецепторы (PRR), альвеолярные эпителиальные клетки и альвеолярные макрофаги обнаруживают высвобожденные патоген-ассоциированные молекулярные структуры (PAMP), такие как вирусная РНК, и связанные с повреждением молекулярные структуры (DAMP), включая АТФ, ДНК и ASC олигомеры. Возникает волна локального воспаления, стимулирующая повышенную секрецию провоспалительных цитокинов и хемокинов IL-6, IFNγ, MCP1 и IP-10 в кровь [11, 22]. Эти цитокины являются индикаторами поляризованного ответа T-хелперов 1 типа (Тх1), что аналогично наблюдениям, проведенным для SARS-CoV и MERS-CoV [56]. Секреция таких цитокинов и хемокинов привлекает иммунные клетки, в частности моноциты и Т-лимфоциты, но не нейтрофилы, из крови в очаг инфекции [57, 58]. Рекрутирование иммунных клеток из кровотока в легкие и инфильтрация дыхательных путей лимфоцитами может объяснить лимфопению и увеличение соотношения нейтрофилов и лимфоцитов, наблюдаемого примерно у 80 % пациентов с инфекцией SARS-CoV-2 [6, 59].
У большинства людей рекрутированные клетки элиминируют инфекцию в легких, иммунный ответ уменьшается, и пациенты выздоравливают. Однако у некоторых пациентов возникает дисфункциональный иммунный ответ, который запускает цитокиновый шторм, опосредованно вызывающий выраженное воспаление в ткани легких. Было отмечено, что у пациентов с тяжелой формой COVID-19, нуждающихся в интенсивной терапии в больницах, наблюдались более высокие уровни IL-2, IL-7, IL-10, гранулоцитарного колониестимулирующего фактора (G-CSF), IP-10, MCP1, макрофагального воспалительного белка 1α (MIP1α) и фактора некроза опухоли (ФНО) в плазме крови [11]. Уровни IL-6 у этих пациентов продолжали увеличиваться с течением времени и являлись относительно более высокими у погибших, чем у выживших [60].
Примечательно, что в жидкости бронхоальвеолярного лаважа у пациентов с тяжелым течением COVID-19 определяется обширная воспалительная моноцитарная популяция макрофагов FCN1+ [61]. Кроме того, у пациентов с тяжелым заболеванием значительно выше процент воспалительных моноцитов CD14+ и CD16+ в периферической крови, чем у пациентов с легким течением болезни [62]. Эти клетки секретируют воспалительные цитокины, включая MCP1, IP-10 и MIP1α, которые способствуют развитию цитокинового шторма (рис. 1).
Механизмы, с помощью которых SARS-CoV-2 подрывает врожденные функции противовирусных цитокинов, еще предстоит изучить, но исследование SARS-CoV показывает, что множественные структурные и неструктурные белки вируса противодействуют интерфероновому ответу. Антагонизм возникает на различных этапах передачи сигналов интерферона, в том числе путем предотвращения распознавания паттерн-распознающим рецептором (PRR) вирусной РНК [63-65], путем предотвращения передачи сигналов PRR через TBK1 / ингибитор субъединицы ε киназы транскрипционного фактора NF-κB (каппа-би) (IKKε), TRAF3 и IRF3 [63, 66], предотвращая передачу сигналов интерферона далее через STAT1 [67] и способствуя деградации мРНК хозяина, ингибируя трансляцию белка [68]. Весьма вероятно, что по крайней мере некоторые из этих механизмов аналогичны и для SARS-CoV-2. Антагонизм белков вируса по отношению к интерфероновому ответу способствует репликации вируса, что приводит к увеличению высвобождения продуктов пироптоза, которые могут дополнительно вызывать аберрантные воспалительные реакции.
Неограниченная воспалительная инфильтрация клеток сама по себе может привести к повреждению легких через чрезмерную секрецию протеаз и активных форм кислорода в дополнение к прямому повреждению, вызванному вирусом. Вместе они приводят к диффузному альвеолярному повреждению, включая десквамацию альвеолярных клеток, образование гиалиновых мембран и отек легких [57, 58]. Это ограничивает эффективность газообмена в легких, вызывая затруднение дыхания и приводя к снижению уровня кислорода в крови. Легкие также становятся более уязвимыми для вторичных инфекций.
Вдобавок к локальному повреждению, цитокиновый шторм производит волновые эффекты по всему телу. Повышенные уровни цитокинов, таких как ФНО, могут вызывать септический шок и полиорганную недостаточность. Это может привести к повреждению миокарда и нарушению кровообращения, наблюдаемым у некоторых пациентов [69]. Пожилые пациенты (в возрасте старше 60 лет) и люди с сопутствующими заболеваниями более склонны к развитию такого дисфункционального иммунного ответа, который способствует развитию патологии, а также не позволяет уничтожить возбудителя. Точные причины этого неясны, хотя одной из причин может быть старение микросреды легких, вызывающее измененное созревание дендритных клеток, миграцию в лимфоидные органы [70] и, следовательно, дефектную активацию Т-клеток. Напротив, дети, как правило, не склонны к развитию тяжелой формы заболевания, несмотря на то, что у них могут наблюдаться высокие вирусные титры [71]. Во всех возрастных группах моложе 18 лет более 50 % детей испытывали легкие симптомы или не имели симптомов вовсе, у менее чем 6 % детей развивались тяжелые симптомы [72]. Таким образом, в то время как вышеупомянутые исследования описывают важные достижения, полная картина критических иммунных факторов, которые лежат в основе развития более тяжелых воспалительных реакций у некоторых пациентов, неясна.
Вопрос о том, необходимо ли сохранение вируса для нанесения постоянного ущерба, остается спорным. Пик вирусных титров в образцах клеток дыхательных путей при инфекциях SARS-CoV и SARS-CoV-2 может наблюдаться еще до появления симптомов пневмонии [17, 19]. Тем не менее большое ретроспективное когортное исследование показало, что вирусная РНК обнаруживалась у не выживших пациентов вплоть до момента смерти, что позволяет предположить корреляцию между устойчивостью вируса и неблагоприятным исходом заболевания [60]. Поскольку вирусная РНК может задерживаться даже после активной инфекции и не является показателем инфекционности вируса, вопрос о том, связан ли исход с количеством инфекционных частиц, дискуссионен. Кроме того, более ранние исследования SARS-CoV показали, что вирус, кроме клеток легких, может инфицировать и другие мишени. Примечательно, что вирус был обнаружен в Т-лимфоцитах [73], макрофагах [74–76] и дендритных клетках, происходящих из моноцитов [77]. Непосредственное уничтожение вируса лимфоцитами может способствовать возникновению лимфопении, наблюдаемой у пациентов [73]. Вирусная инфекция в иммунных клетках, таких как моноциты и макрофаги, может привести к аберрантной выработке цитокинов, даже если вирус активно не размножается [74–77]. Степень тропности SARS-CoV-2 к этим клеткам остается неясной. Понимание точных причин иммунной дисфункции имеет решающее значение для применения соответствующих иммуномодулирующих методов лечения.
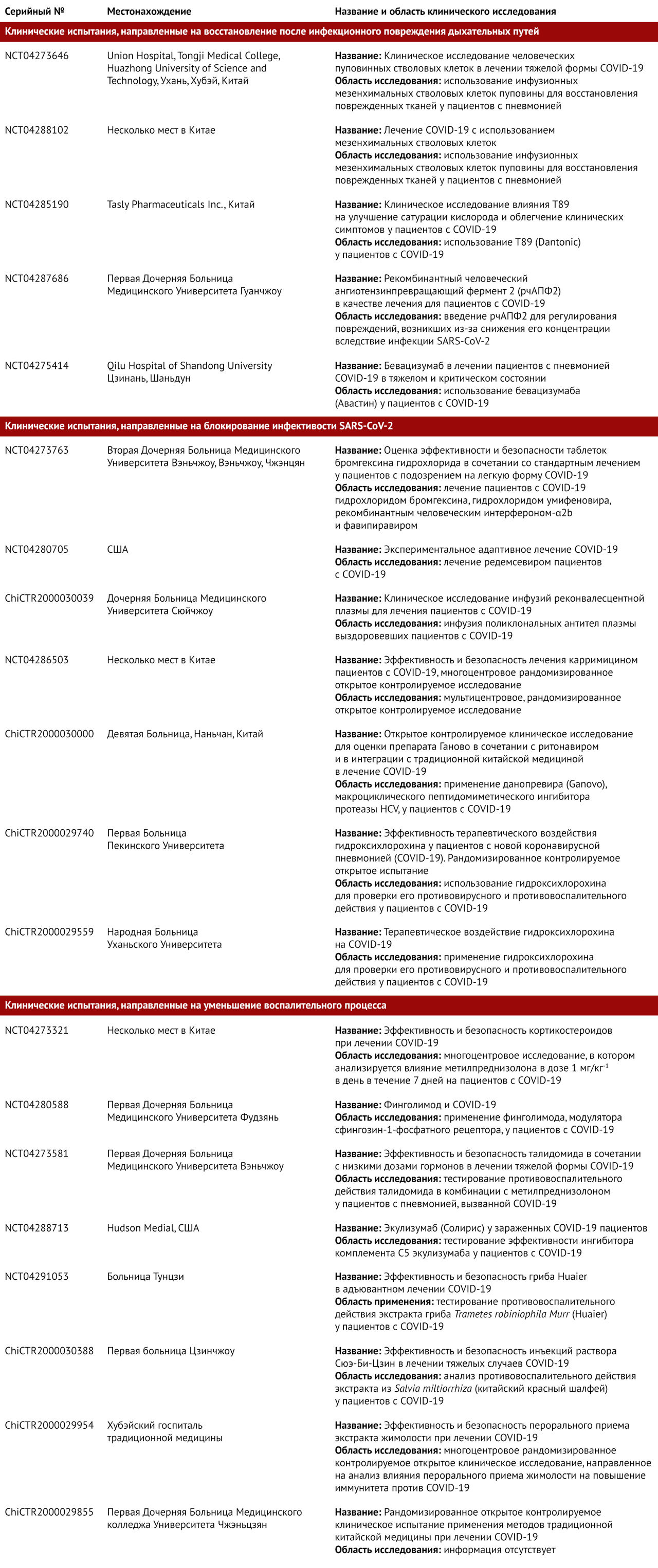
Несколько иммуносупрессивных методик лечения, направленных на ограничение иммуноопосредованого повреждения при COVID-19, находятся на различных этапах разработки и перечислены в таблице 1. В настоящее время проводятся испытания кортикостероидов для лечения COVID-19 [78], хотя этот класс препаратов не рекомендовался во время эпидемии SARS-CoV 2003 г. [79, 80]. Кроме того, проводятся клинические испытания антагониста IL-6 тоцилизумаба для проверки его эффективности [81], а также исследуется сарилумаб [82]. В других клинических испытаниях тестируются эффекты воздействия на колониестимулирующий фактор гранулоцитов и макрофагов (GM-CSF), например, с помощью гимсилумаба [83], лензилумаба [84] и намилумаба [85]. Еще одним новым методом вспомогательной терапии является цитосорб [86], который поглощает разнообразные цитокины, DAMP и PAMPs с целью снижения их уровня в крови и смягчения иммунопатологических процессов.
{kind=link}
Талидомид, препарат с иммуномодулирующими свойствами, также был успешно применен у одного пациента с COVID-19 [87]. В результате на сегодняшний день было начато два клинических испытания, чтобы проверить его потенциал для уменьшения повреждения легких [88, 89]. В контексте инфекции SARS-CoV предлагался, но не был проверен антагонист ФНО, впрочем он также не был проверен на пациентах с COVID-19 [90]. Небольшое открытое нерандомизированное исследование показало, что комбинация гидроксихлорохина (известного противомалярийного средства) и азитромицина (распространенного антибиотика) может оказаться эффективной для лечения пациентов с тяжелой формой COVID-19 [91]. Хотя влияние гидроксихлорохина на прямое ингибирование вируса [92] и его противовоспалительная и иммуномодулирующая активность известны [93], еще предстоит определить, действуют ли эти механизмы в отношении COVID-19 [94].
Т-клеточный иммунитет
Т-клеточный и В-клеточный иммунный ответ на SARS-CoV-2 реализуется примерно через 1 неделю после появления симптомов COVID-19. CD8+ T-клетки важны для прямой атаки и уничтожения инфицированных вирусом клеток, тогда как CD4+ T-клетки имеют решающее значение для активации как CD8+ T-клеток, так и B-клеток. CD4+ T-клетки также ответственны за выработку цитокинов, которые управляют рекрутментом иммунных клеток. Первое вскрытие пациента с COVID-19 выявило накопление мононуклеарных клеток (вероятно, моноцитов и Т-клеток) в легких в сочетании с низким уровнем гиперактивных Т-клеток в периферической крови [57]. Наряду с сообщениями о лимфопении и снижении уровня периферических Т-клеток у пациентов [6, 95–97], эти данные свидетельствуют о том, что Т-клетки привлекаются из кровотока в очаг инфекции для контроля вирусной инфекции. У пациентов с COVID-19 интенсивное истощение Т-клеток и снижение функционального разнообразия указывали на развитие тяжелого течения болезни [98]. Несмотря на ослабленный ответ у пациентов, которые выздоровели от инфекции SARS-CoV, развились специфические для коронавируса Т-клетки памяти, которые были обнаружены через 2 года после выздоровления [99, 100].
SARS-CoV-специфичные CD4+ T-клетки экспрессируют IFNγ, ФНО и IL-2, что позволяет предположить, что у пациентов с инфекцией SARS-CoV проявляется Тх1-клеточный ответ, и для контроля инфекции используется в основном клеточный иммунитет [101, 102]. Хотя такой провоспалительный профиль может быть фактором, ухудшающим иммунопатогенез, было предположено что T-клетки CD4+ угнетают SARS, поскольку истощение пула этих клеток у мышей приводило к более медленной элиминации вируса из клетки хозяина и более выраженному воспалению легких [103]. При использовании адаптированного для мышей штамма SARS-CoV иммунизация дендритными клетками, несущими пептиды SARS-CoV, привела к увеличению числа вирус-специфических CD4+ и CD8+ T-клеток, которые накапливались в легких и способствовали увеличению выживаемости [104, 105]. Кроме того, введение SARS-CoV-специфических CD4+ и CD8+ T-клеток иммунодефицитным мышам приводило к улучшению защиты против адаптированного для мышей штамма SARS-CoV [105].
Несмотря на доказательства важной роли Т-клеток в борьбе с инфекцией, несколько вакцин против SARS-CoV, ранее протестированных на животных моделях, выявили признаки иммунопатологии, связанной с Тх2-опосредованной эозинофильной инфильтрацией [106,107]. В частности, у старых мышей, которые были вакцинированы, наблюдалось скорее усугубление иммунной патологии, а не защита [108]. Дальнейшее изучение природы защитного и дисфункционального Т-клеточного ответа необходимо для определения оптимальных стратегий использования Т-клеток в разработке вакцин [109]. Коронавирус-специфические Т-клетки, несомненно, важны для устранения вируса и контроля развития заболевания и должны учитываться в стратегиях вакцинации. Однако вопрос о том, способен ли Т-клеточный иммунный ответ предотвращать инфекцию у людей, еще предстоит прояснить. Эти знания будут важны для разработки вакцин.
В-клеточный (гуморальный) иммунитет
B-клеточный иммунный ответ у пациентов с COVID-19 возникает одновременно с реакцией T-хелперных фолликулярных клеток примерно через 1 неделю после появления симптомов [110]. У пациентов с инфекцией SARS-CoV B-клеточный ответ обычно возникает в первую очередь против белка нуклеокапсида (N). В течение 4–8 дней после появления первых симптомов обнаруживается повышение титра антител к шиповидным белкам [111, 112]. Нейтрализующие антитела к шиповидным белкам начинают вырабатываться ко 2 неделе заболевания, а у большинства пациентов нейтрализующие антитела вырабатываются к 3 неделе [113, 114]. Учитывая, что вирусная нагрузка достигает пика для SARS-CoV-2 раньше, чем для SARS-CoV [16–19], выработка антител также может начинаться раньше. Похоже, что у некоторой части пациентов долгосрочные антитела к SARS-CoV-2 могут не вырабатываться [115]. Остается неизвестным, подвержены ли эти пациенты повторному заражению, о котором есть отдельные сообщения [116, 117].
Вероятно, против SARS-CoV-2 будут эффективны антитела: образцы реконвалесцентной плазмы были применены в отношении COVID-19 с хорошими клиническими результатами [118], также они ранее успешно использовались при лечении ОРДС [119–121].
Хотя механистические корреляты защиты у людей еще не идентифицированы, считается, что нейтрализация вируса является одной из важных функций антител, хотя конкретный титр и специфичность требуемого для защиты набора антител остаются неопределенными. В SARS-CoV основной мишенью нейтрализующих антител является RBD [122], содержащий 193 аминокислотных остатка (аминокислоты 318–510) в шиповидном белке, которые могут независимо связываться с рецептором АПФ2 — мишенью клетки-хозяина [35–37]. Хотя некоторые из ранее идентифицированных моноклональных антител к SARS-CoV также связываются и с SARS-CoV-2 [123], большинство все же не делают этого [124], что может быть связано со значительными различиями в RBD SARS-CoV-2 и SARS-CoV (рис. 4). В частности, из 33 аминокислот в регионе 460–492 аминокислоты шиповидного белка SARS-CoV, содержащего ключевые остатки, связывающиеся с АПФ2 [125], менее половины (15/33) сохраняются в SARS-CoV-2. Тем не менее мышиная антисыворотка против белка SARS-CoV может перекрестно нейтрализовать псевдовирус SARS-CoV-2, что указывает на перекрывающиеся нейтрализующие эпитопы между двумя вирусами [28, 126].
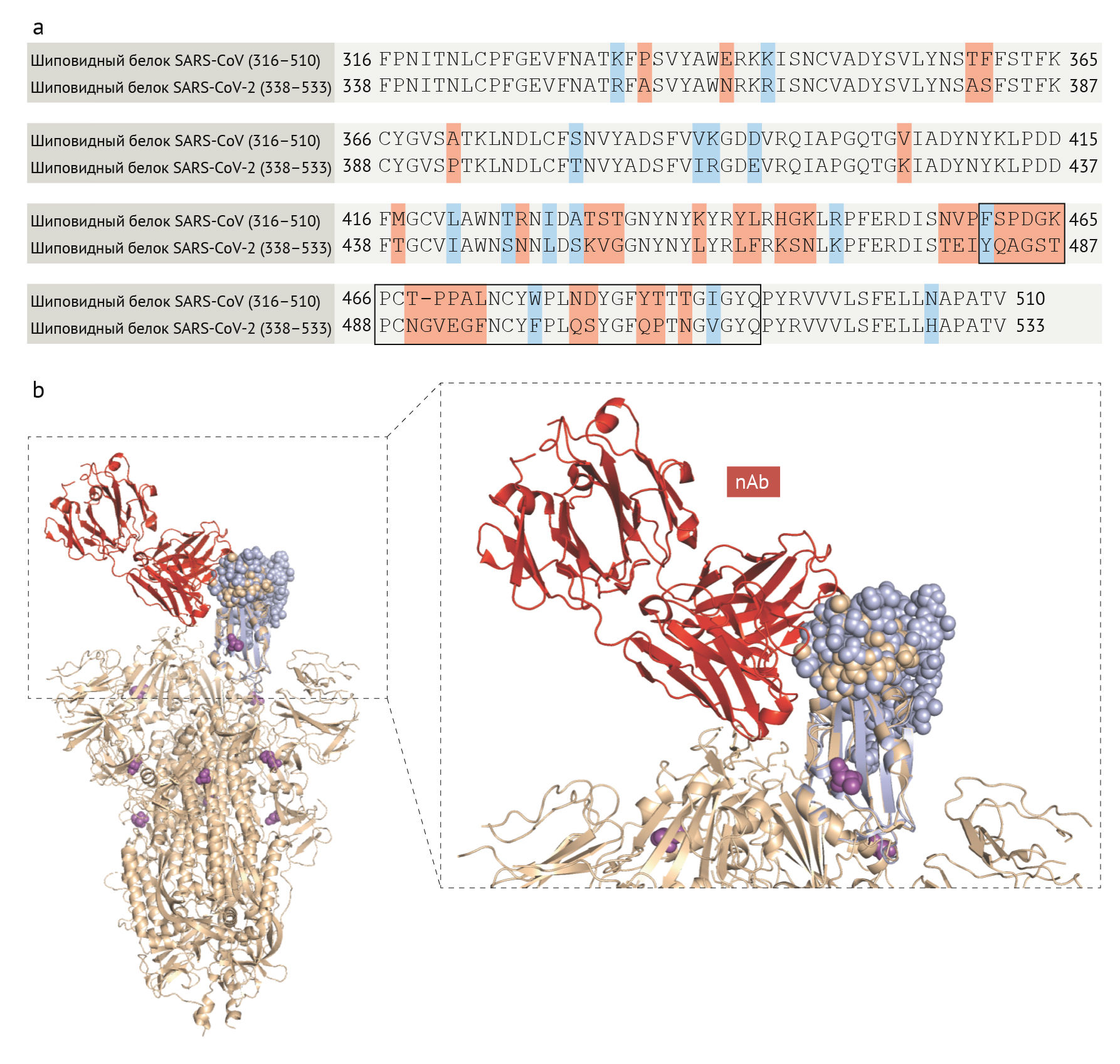
(b) 3D-структура SARS-CoV-2 (Protein Data Bank ID 6VSB42, персиковая лента) наложена на SARS-CoV рецептор-связывающий мотив (RBM) с нейтрализующим антителом (nAb; красная лента), взаимодействующим с RBM (протеином). Банк данных 2DD8 [151], фиолетовая лента). Персиковая и фиолетовая сферы обозначают RBM SARS-CoV-2 и SARS-CoV соответственно. Пурпурные сферы обозначают несинонимические изменения в шиповидном белке SARS-CoV-2, о которых сообщалось ранее [135].
В Китае больницы начали использовать плазму реконвалесцентов в качестве источника поликлональных антител для лечения COVID-19, и ранние данные свидетельствуют о положительном влиянии в виде снижения вирусной нагрузки и смертности [127, 128]. В настоящее время предпринимаются усилия для разработки терапевтических моноклональных антител против SARS-CoV-2 с использованием таких подходов, как отображение фаговой библиотеки, традиционная иммунизация мыши и создание гибридомы, а также клонирование B-клеточных последовательностей людей-реконвалесцентов [129–132]. SARS-CoV, по-видимому, не имеет механизмов для предотвращения нейтрализации антителами, таких как экранирование гликаном сайта связывания рецептора антителом [133]. Это дополнительно подтверждается тем фактом, что организм пациентов с инфекцией SARS-CoV, как правило, был способен вырабатывать нейтрализующие антитела. Фрагмент рекомбинантного шиповидного белка, который включал в себя RBD SARS-CoV, показал самую высокую иммунногенность по сравнению с другими протестированными фрагментами рекомбинантного шиповидного белка, что позволяет предположить, что иммунная система способна эффективно нацеливаться на нейтрализующие эпитопы [134]. Таким образом, если SARS-CoV-2 ведет себя так же, как и SARS-CoV в этом отношении, вполне вероятно, что усилия по разработке нейтрализующих моноклональных антител будут оправданы.
Возможно, изменения в шиповидном белке сделают SARS-CoV-2 устойчивым к некоторым моноклональным антителам, особенно когда он мутирует. На данный момент весь RBD остается неизменным, и существует только четыре известных редких несинонимических изменения в шиповидном белке: V483A, L455I, F456V и G476S135. Изменение V483A сопоставляется с аналогичным естественным изменением, обнаруженным в MERS-CoV, I529T, где оно снижает связывание вирусного белка с рецептором-мишенью хозяина, а также повышает устойчивость к нейтрализации антителами из образцов сыворотки от пациентов с MERS [136]. Изменения F456V и G476S также соответствуют сходным позициям изменения у SARS-CoV (L443R и D463G), которые были обнаружены на панели избежавших нейтрализации мутантных вирусных частиц [137].
Так или иначе выбор кандидатов для терапии антителами должен включать тщательное рассмотрение возможных нежелательных побочных эффектов. Например, ранее существующие антитела к другим коронавирусам могут обострять инфекцию SARS-CoV посредством антителозависимого усиления [138–140]. Кроме того, предыдущие исследования на животных показали, что при инфекции SARS-CoV нейтрализирующие антитела к шиповидным белкам могут потенциально усиливать тяжелое повреждение легких, усугубляя воспалительные реакции [141]. Также наблюдалась корреляция между развитием ОРДС и сероконверсией противовирусного IgG у 80 % пациентов [19]. У пациентов, организм которых выработал нейтрализирующие антитела к шиповидным белкам ранее, заболеваемость была выше; пациентам, умершим от инфекции, потребовалось в среднем всего 14,7 дня, чтобы достичь пиковых уровней активности нейтрализующих антител, в отличие от 20 дней у пациентов, которые выздоравливали [142]. Аналогично для MERS: у пациентов с более тяжелым течением заболевания титры антител, по-видимому, выше, чем у пациентов с легким заболеванием [143, 144], хотя в одном исследовании подвергается сомнению тот факт, что задержка в выработке антител связана с заболеванием [145]. Связывание иммунных комплексов антитело-вирус с активирующими Fc-рецепторами на альвеолярных макрофагах может индуцировать экспрессию провоспалительных факторов, включая IL-8 и MCP1, которые присоединяются к иммуностимулирующей среде [146]. Такие комплексы могут также активировать систему комплемента и приводить к дальнейшему нежелательному воспалению [141]. Таким образом, важно рассмотреть вопрос о разработке терапевтических антител, обладающих низкой или отсутствующей провоспалительной активностью, но сохраняющих свою способность нейтрализовать вирусы [147]. Например, могут быть внесены изменения в области Fc и/или проведено его гликозилирование, чтобы изменить аффинность связывания для активации рецепторов Fc [146, 148].
Выводы
В этом обзоре представлены различные патофизиологические механизмы инфекции SARS-CoV-2 и иммунопатогенеза COVID-19. Контроль воспалительного ответа может быть столь же важным, как таргетированное воздействие на вирус. Препараты, ингибирующие вирусную инфекцию и регулирующие дисфункциональные иммунные реакции, могут быть использованы совместно для борьбы с патологией в несколько этапов. В то же время связь между иммунной дисфункцией и тяжестью исхода заболевания у пациентов с COVID-19 должна служить предостережением при разработке и оценке вакцины. Необходимы дальнейшие исследования иммунного ответа хозяина на SARS-CoV-2, в том числе подробное изучение детерминант благоприятных и дисфункциональных исходов. Они также помогут идентифицировать биомаркеры для определения иммунных коррелятов защиты и тяжести заболевания для эффективной сортировки пациентов.
