
Критический период онтогенеза, определяющий формирование нейрональных двигательных цепей
У плодовых мушек был обнаружен механизм, который позволяет клеткам, называемым астроцитами, передавать сигналы нейронам, что закрывает пробел в понимании эмбриологического этапа, в течение которого формируется паттерн воспроизведения локомоторных актов.
В развитии организма существуют периоды, когда части формирующейся нервной системы особенно чувствительны к изменяющимся воздействиям внешней среды. Нарушение этих критических периодов может иметь жизненно важные последствия для нейронных связей и функционирования нервной ткани мозга [1]. Например, для периода детства критическим является момент овладения языком [2]. Было высказано предположение, что изменение хода развития организма в течение критических периодов в определенной степени отражается в патологиях развития нервной системы (к примеру, расстройстве аутистического спектра [3] и шизофрении [4]). Критические периоды подробно описаны применительно к патологиям зрительной системы [1], но до сих пор системам, не относящимся к группе сенсорных, уделялось меньше внимания. Своей статьей в «Nature» Ackerman с соавт. [5] закрывают этот пробел. Исследователями определен критический период для развития двигательных цепей у плодовых мушек Drosophila melanogaster и установлены клеточные и молекулярные основы завершения критического периода в формировании системы локомоции.
Существует несколько способов изменения нейронных связей на протяжении критического периода. Ackerman с соавт. в основном касаются гомеостатической пластичности, при которой изменения происходят во всем нейроне, включая размеры структур, называемых дендритами, которые получают синаптические связи от других нейронов; количество синапсов; силу электрических импульсов, передаваемых синапсами [6].
В первую очередь, авторы воспользовались методикой, называемой оптогенетикой, для активации или подавления нейрональной активности в двух классах нервных клеток, называемых мотонейронами aCC и RP2. Когда активность нейронов приглушалась, длина и объем дендритов клеток увеличивались. Напротив, оптогенетическая активация приводила к ретракции дендритов. Эти изменения происходили только тогда, когда манипуляции с нейронной активностью осуществлялись в течение восьми часов после вылупления личинок, и всего 15 минут хватало, чтобы увидеть эффекты.
Затем Ackerman с соавт. задались вопросом, отражается ли изменение формы дендритов на изменении количества возбуждающих и тормозных синапсов мотонейронов aCC и RP2 (эти синапсы ответственны за активацию и ингибирование активности нейронов, соответственно). Оптогенетическое «приглушение» нейронов привело к снижению числа тормозных синапсов и увеличению количества возбуждающих синапсов. Благодаря обоим процессам — расслаблению дендритов и изменению состава синапсов — равновесие нейрональной активности было восстановлено, что противодействовало эффекту оптогенетического «приглушения». Оптогенетическая активация нейронов aCC и RP2 привела к снижению количества возбуждающих синапсов, однако количество тормозных синапсов не уменьшилось. Возможно, это связано с ограниченным пространством клеточной мембраны, доступной для образования синаптических контактов после ретракции дендритов. Вместе эти выводы указывают на то, что существует критический период для гомеостатических изменений структуры дендритов и числа синапсов в развивающейся двигательной системе D. melanogaster (рис. 1a, b).
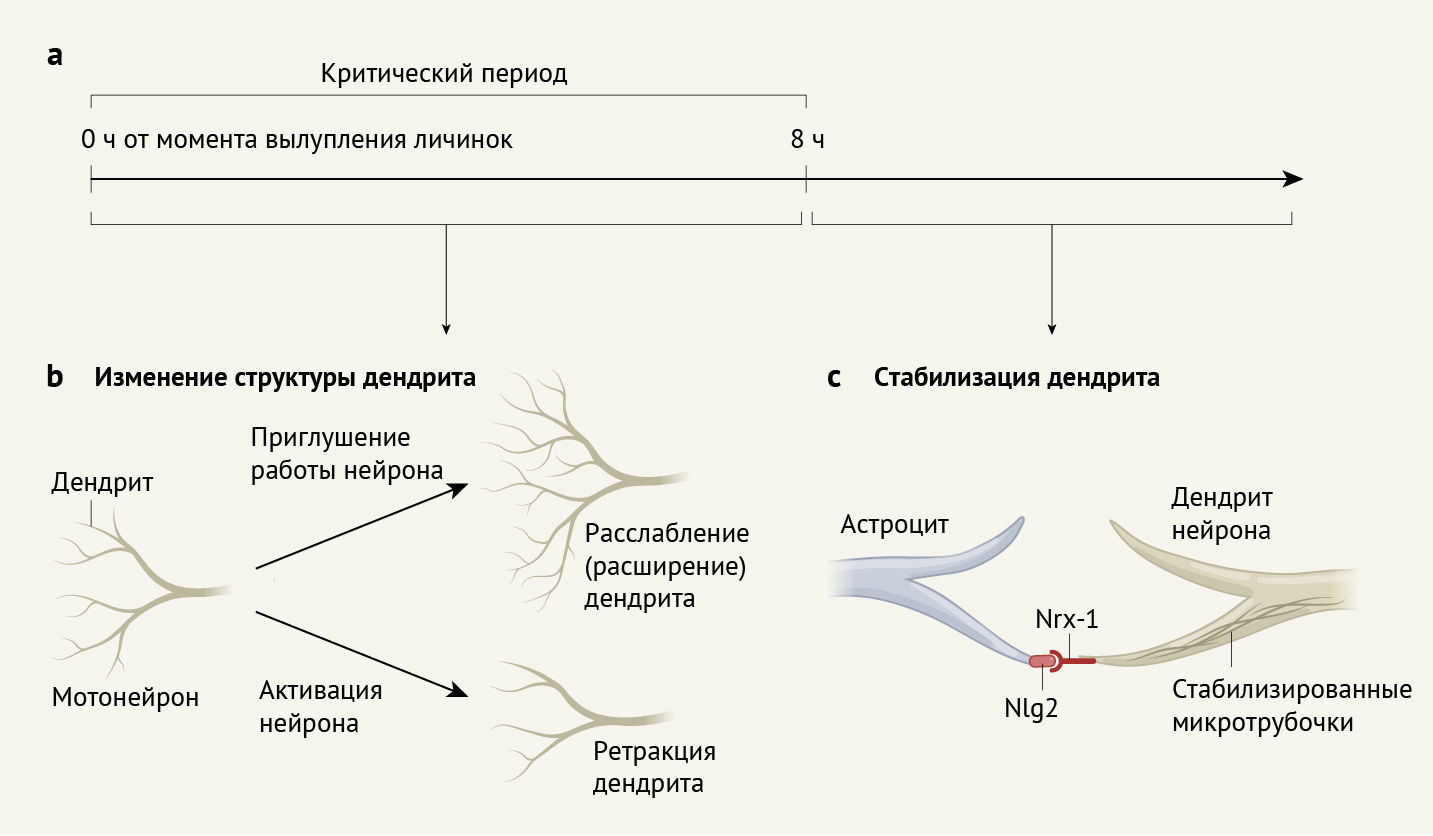
b. Приглушение активности нейронов в критический период приводит к расслаблению дендритов. И наоборот, активация нейронов приводит к ретракции дендритов.
c. Критический период завершается по мере созревания соседних клеток, называемых астроцитами. Созревающие астроциты синтезируют белок Nlg2, который взаимодействует с белком Nrx-1 на дендрите нейронов. Это взаимодействие приводит к стабилизации структур, называемых микротрубочками, что, в свою очередь, предотвращает дальнейшее ремоделирование дендритов.
Что является причиной этих изменений? Часто нейроны находятся в тесном контакте с клетками, называемыми астроцитами, которые помогают регулировать развитие синапсов и поддерживать функционирование ткани мозга [7], поэтому Ackerman c соавт. использовали генную инженерию для уничтожения всех астроцитов у плодовых мушек в эксперименте. Ремоделирование дендритов продолжалось спустя более восьми часов от момента вылупления личинок этих мух-мутантов, однако до истечения периода в восемь часов такового не наблюдалось. Это свидетельствует о том, что астроциты регулируют время завершения критического периода для системы aCC/RP2, а не возможность нейрональной пластичности в это время. Это ключевое различие, поскольку согласно нему, в основе этих описанных явлений лежат разные механизмы.
Следовательно, Ackerman с соавт. стремились определить механизмы, ответственные за завершение критического периода. Учеными использовался метод, называемый скринингом РНК-интерференции, для подавления трансляции белков с различных информационных РНК в астроцитах личинок. Затем было проанализировано время завершения критического периода у каждой из развившихся мух. Это позволило выявить гены, с помощью которых осуществляется завершение критического периода. Как обнаружилось, существует несколько генов, для которых интерференция РНК продлевала критический период, однако во многих случаях ингибирование этих генов также оказывало глубокое влияние на форму астроцитов, затрудняя возможность различения их роли в развитии астроцитов от конкретной роли в регулировании критических периодов. Авторы решили сосредоточиться на гене nlg2, ингибирование которого продлевает критический период без изменения формы астроцитов.
Эквивалентное семейство генов у мышей, именуемое нейролигинами, было связано с созреванием астроцитов во время критического периода в зрительной коре головного мозга, и созревание астроцитов близко совпадает с завершением этого критического периода [8]. У D. melanogaster белок Nlg2 взаимодействует с белком neurexin-1 (Nrx-1), который был обнаружен Ackerman с соавт. в дендритах мотонейронов. Авторам удалось показать, что ингибирование nrx-1 в нейронах aCC и RP2 с использованием интерференции РНК увеличивает критический период. Таким образом, Nrx-1, вероятно, является нейрональным рецептором для астроцитарного Nlg2 при регулировании завершения критического периода (рис. 1c). В соответствии с этой идеей, избыточная экспрессия либо nlg2 в астроцитах, либо nrx-1 в дендритах нейронов типа aCC или RP2 преждевременно прекращала критический период, сокращая его до четырех часов от момента вылупления личинок.
Нарушение критических периодов может иметь длительные последствия для функционирования нервных цепей. В подтверждение этому Ackerman с соавт. обнаружили, что продление критического периода (посредством воздействия либо на nrx-1, либо на nlg2) приводит к аномальному двигательному поведению. При этом возникновение паттерна движений личинки в виде аномальной спирали через полтора дня после проведения процедуры является хорошим периодом времени для анализа. На этой стадии личинки активно питаются и перемещаются по питательной среде. Эти изменения в поведении подчеркивают важность правильного выбора времени для критических периодов.
Работа Ackerman с соавт. поднимает новые вопросы для будущих экспериментов. Например, как взаимодействие между Nlg2 и Nrx-1 определяет протекание критического периода? Настоящей работой была определена роль этого взаимодействия в стабилизации структурных полимеров, называемых микротрубочками, и, таким образом, в стабилизации структуры самих дендритов. Однако механизм, лежащий в основе стабильности микротрубочек, еще предстоит изучить. Кроме того, еще предстоит определить динамику синтезирования Nlg2 и Nrx-1. Могут ли эти белки регулироваться ретракцией или расслаблением дендритов? Окажется ли ретракция дендритов результатом снижения экспрессии nlg2 в астроцитах?
Авторами исследования предоставлены доказательства важной роли астроцитов в регуляции завершения критического периода. Астроциты регулируют аспекты пластичности критических периодов в зрительной системе млекопитающих [9], в том числе посредством воздействия секретируемыми хордино 1- и хевино-подобными белками [10–11]. В настоящей работе показано, что астроциты могут регулировать критические периоды не только в сенсорных, но и в двигательных системах. Выявление механизмов, управляющих течением критических периодов, представляет особый интерес, потому что изменения в их ходе могут нарушить правильное развитие нервной системы. Следовательно, полученные данные могут привести к пониманию механизмов, задействованных в нарушениях психического развития, таких как шизофрения [3]. Кроме того, выявление механизмов завершения критических периодов может позволить лучше разобраться, как свойство пластичности нервной ткани мозга утрачивается во взрослом возрасте, что открывает новые возможности для терапевтического воздействия, направленного на повышение этой характеристики после травмы или болезни мозга.
Работа также показывает, что роль астроцитов в регулировании критических периодов присутствует и у беспозвоночных, что подчеркивает центральную роль этих клеток в развитии и созревании нервной системы. Становится очевидным, что астроциты и связанные с ними клетки (вместе называемые нейроглией) являются главными регуляторами пластичности нейронов, особенно в контексте гомеостатических изменений и изменений цепей взаимосвязей нейронов. Последующие исследования критических периодов должны учитывать важную роль глии.
