
Биосинтез макромолекул

В основе жизнедеятельности клетки лежит процесс интерпретации данных с дезоксирибонуклеиновой кислоты (ДНК). Но прежде чем показать, как все это происходит, я напомню базовые моменты.
▶ Материал по теме: Подкаст о биохимии (и зачем она врачу)
Введение
ДНК — это полимер, состоящий из дезоксирибонуклеотидов — нуклеотидов, у которых «отобрали» ОН-группу при втором атоме рибозы. Получается это в ходе следующей реакции (рис. 1):
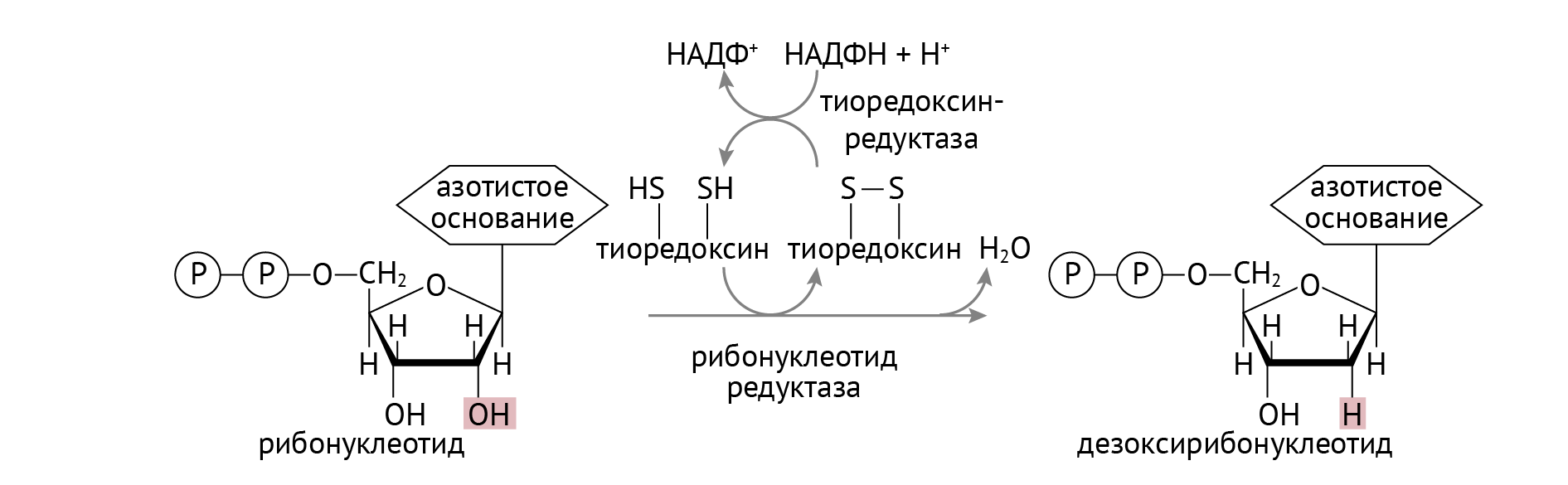
Рисунок 1 | Образование дезоксирибонуклеотидов
Стоит обратить внимание на важную роль НАДФН, который участвует в восстановлении тиоредоксина в тиоредоксинредуктазной реакции (см. рис. 1). Источником НАДФН является пентозофосфатный цикл глюкозы, о котором я расскажу в следующей статье. В отличие от РНК, в состав ДНК входят нуклеотиды с тимином, а не с урацилом. Откуда берется тимин, показано в реакции, изображенной на рис. 2.
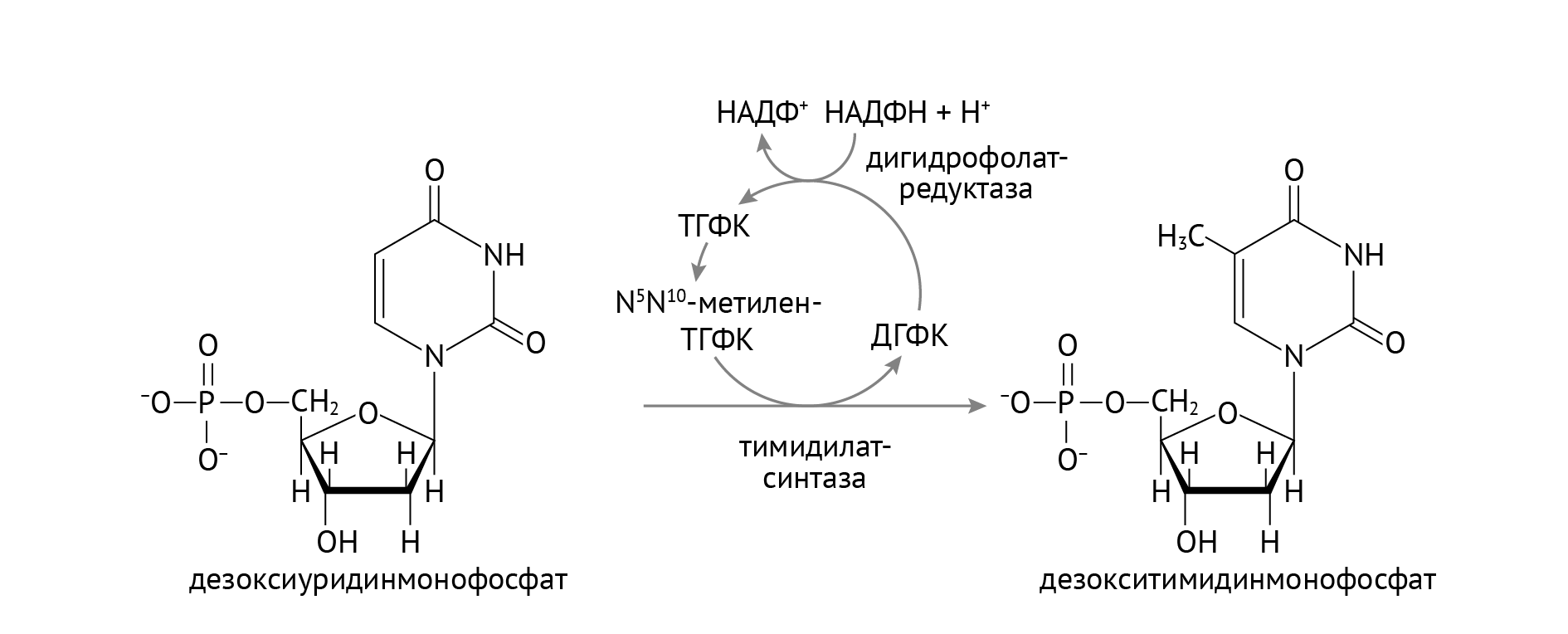
Рисунок 2 | Образование дезокситимидинмонофосфата
Здесь также большое значение имеет НАДФН из пентозофосфатного цикла. Однако помимо НАДФН, важными участниками образования тимина (если быть занудой, дезокситимидинмонофосфата) являются фолиевая кислота и витамин В12 (восстанавливает фолиевую кислоту до активной формы, не указан на данном рисунке) [1]. Это объясняет механизм мегалобластной анемии, а также именно эта реакция является фармакологической мишенью таких противоопухолевых препаратов, как 5-фторурацил и метотрексат.
Субстратом для синтеза нуклеиновых кислот, как известно из прошлой статьи, служат пуриновые (аденин, гуанин, которые часто реутилизируются) и пиримидиновые (урацил, цитозин, тимин) нуклеотиды. Образование азотистых оснований требует активного вовлечения аминокислот (аспартат, глутамин, глицин), фолиевой кислоты и даже аммиака (в виде NH4+ иона). Нуклеотиды синтезируются эндогенно, и биохимическую цепочку превращений здесь можно представить в упрощенном виде следующим образом: аминокислоты → азотистые основания → нуклеотиды → нуклеиновые кислоты. Поэтому белковая недостаточность (ситуация, когда белков и аминокислот в организме расходуется больше, чем поступает) может привести к ухудшению пролиферативных и регенераторных процессов [1, 3, 8].
Структура ДНК
Уровни структуры ДНК
Для понимания того, что далее будет описано в этой статье, тебе следует помнить об уровнях структуры ДНК. Материал для меня самого очень непростой, но фундаментальный, и потому крайне важный.
Первичная структура ДНК — последовательность дезоксирибонуклеотидов (ДРН) в виде единой полинуклеотидной цепи.
Вторичная структура ДНК — это две полинуклеотидные цепи, соединенные между собой водородными связями и упакованные в виде спирали.
В ходе последовательной укладки при участии гистоновых белков формируется третичная структура ДНК, которую называют хромосомой. В клетке она располагается в комплексе с белками (нуклеопротеидами), и весь этот комплекс (хромосома + белки) носит название хроматина. Чуть позже мы рассмотрим, каковы современные представления относительно пространственной организации хроматина в клетке, и выясним, почему здесь все не так просто, как говорят в университете и пишут в учебниках.
Функциональные элементы хромосом. Что вообще можно найти в нашем геноме?
На рис. 3 схематически представлены функциональные составляющие нашего генома. Можно обратить внимание на то, что гены — участки ДНК, кодирующие информацию о структуре различных полипептидов, составляют всего 30 % от всего генома. Если посмотреть еще внимательнее, то на схеме можно найти такое слово, как экзоны — последовательности ДНК, которые несут информацию о структуре полипептидов и РНК. Удивительно, но получается, что лишь 1,5 % нашего генома что-либо кодирует. А зачем тогда вообще остальные составляющие? Попробуем разобраться и не сойти с ума. Начнем по порядку.
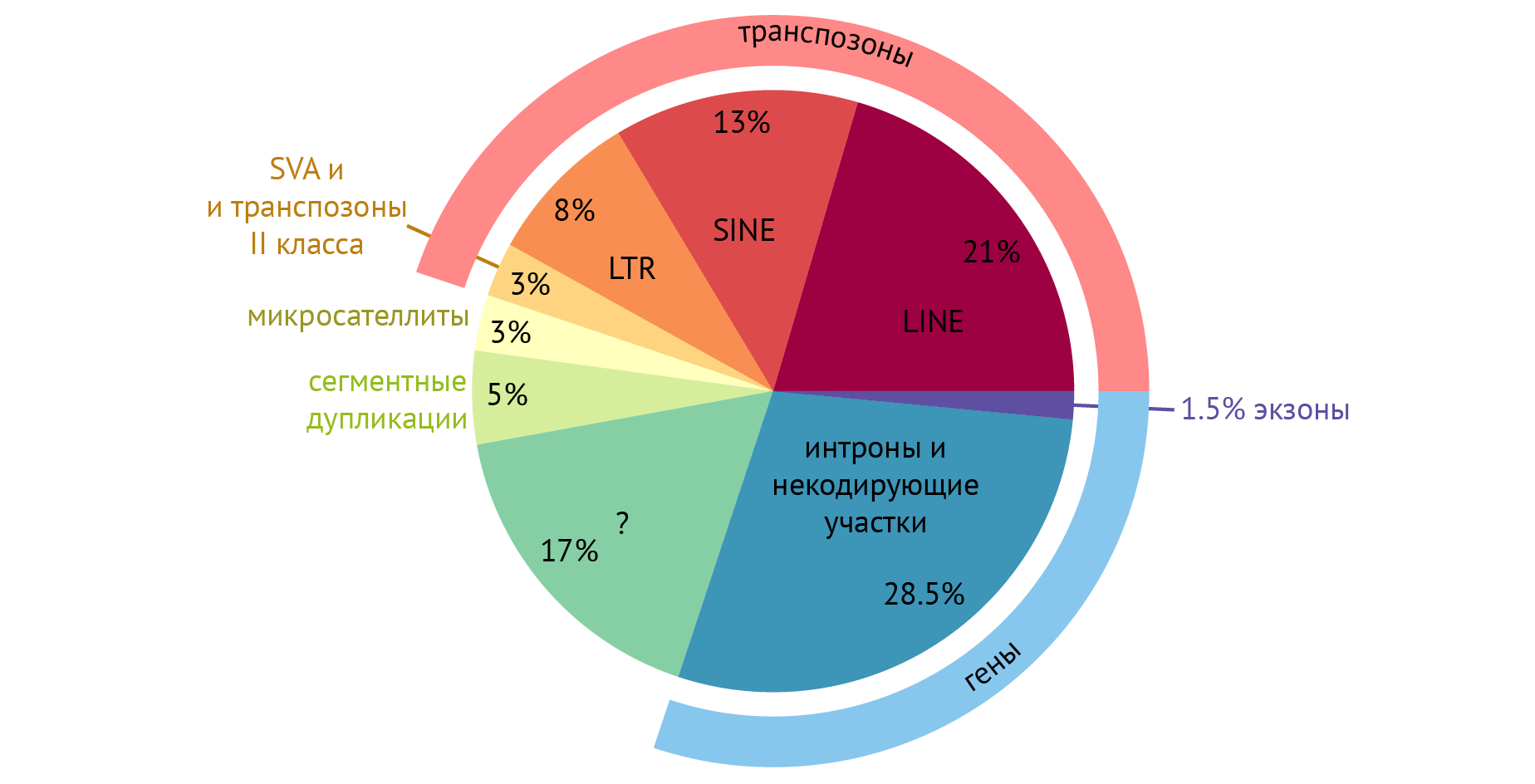
Рисунок 3 | Типы последовательностей в геноме человека
Гены и догма молекулярной биологии
Ген — участок ДНК, несущий информацию о функциональным полипептиде или о молекуле РНК. Ген кодирует первичную последовательность аминокислот.
Согласно центральной догме молекулярной биологии (которая, кстати, уже давно не догма), биологическая информация, заключенная в геноме, превращается в функционально активные белки путем серии процессов (ранее считалось, что они однонаправлены), которые можно схематично изобразить так:
- ДНК → (транскрипция) → РНК → (трансляция) → белок
Нетрудно заметить, что первым процессом является транскрипция — процесс считывания информации об аминокислотной последовательности пептидов с ДНК и перенос этой информации на рибонуклеиновую кислоту (РНК).
Информация о структуре полипептидов заключена именно в генах. Ген в структурном отношении представляет из себя совокупность кодонов — участков ДНК, состоящих из трех нуклеотидов. Каждый кодон кодирует одну аминокислоту: это достаточно простой принцип кодирования [3].
Как уже стало ясно, далеко не все участки ДНК обязательно что-то кодируют… Или нет?
Проект ENCODE
«Около 99 % ДНК человека в принципе ничего не кодирует и не несет какой-либо информации о белках или РНК». Этот тезис разбивается в пух и прах при знакомстве с результатами работы проекта ENCODE, с которыми можно ознакомиться тут. Один из главных выводов заключается в следующем: бо́льшая часть (80,4 %) генома человека участвует по крайней мере в одном биохимическом событии, связанном с РНК и/или хроматином [16].
Если говорить непосредственно о генах, то в них все же встречаются некодирующие последовательности, и называются они интронами. Они к месту или не к месту вставлены внутри генов между кодирующими участками ДНК, которые несут какую-либо информацию и называются экзонами [6]. В генах интроны перемешаны с экзонами. Количество интронов в одном гене может достигать 85 %. К чему такие сложности?
У прокариот лишь немногие гены содержат интроны, а у эукариот интронов очень много. Сравни сложность организации бактерий и сложность организации студента обыкновенного. Еще больше вопросов вызывает тот факт, что генов-то у нас не намного больше.
Считается, что такое усложнение в структуре ДНК является следствием сложности нашей биологической организации: чтобы построить такое произведение биологического искусства, как наш организм, требуется очень тонкая регулировка процессов реализации генетической информации. Один из аспектов этой тонкой регулировки — жонглирование экзонами и интронами на посттранскрипционном этапе, который называется альтернативным сплайсингом. Об этом информация будет чуть ниже, но пока могу сказать, что явление альтернативного сплайсинга позволяет получить множество разнообразных белков из ограниченного числа генов. По-видимому, для достижения биологического разнообразия клеткам проще было научиться управлять малым числом генов, чем увеличивать их количество.
Транспозоны
Транспозоны — подвижные генетические элементы, которые перемещаются с места на место внутри генома по не до конца изученным принципам. Транспозоны составляют до половины нашего генома, и у многих исследователей возникает логичный вопрос: почему? Каково их значение?
На сегодняшний день накоплены данные, свидетельствующие о видообразующей роли транспозонов [14]. Считается, что именно транспозоны, эти активные, подвижные, часто мутирующие элементы ДНК, могут быть одной из причин генетического разнообразия среди разных видов. Между тем, в контексте отдельно взятой бренной человеческой жизни, роль транспозонов сводится к реализации программы онтогенеза, а еще к возникновению различных заболеваний. Связано это с тем, что под действием различных стимулов, в том числе стрессовых для клетки, транспозоны способны менять свое расположение в геноме. В ряде случаев это способно изменить структуру генов и характер их считывания, а следовательно, структуру и функцию кодируемого генами белка. К слову, именно транспозоны являются участками ДНК, которые претерпевают мутации наиболее часто.
Выделяют аж четыре класса транспозонов, но функционально активным считается только один класс — ретротранспозоны, а именно элементы с длинными вкраплениями (LINE1 — TE). Существование транспозонов является одним из опровержений догмы молекулярной биологии, поскольку механизм встраивания транспозона в различные участки генома включает в себя процессы обратной транскрипции (см. рис. 4).
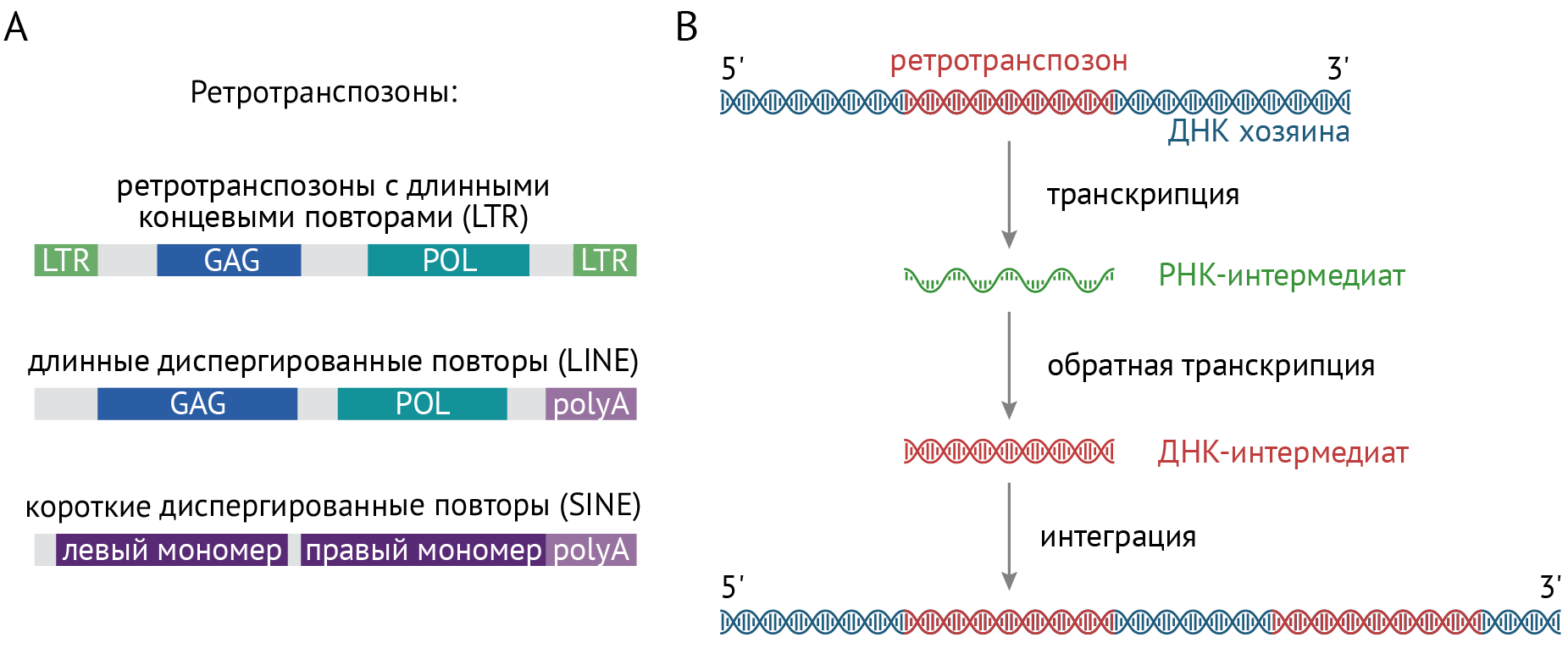
Рисунок 4 | Механизм встраивания транспозона
В результате перемещения транспозонов по геному возможны изменения экспрессии определенных генов, что может вносить свой вклад в развитие различных заболеваний, главным образом опухолевых. Одним из примеров является мутантная вставка (транспозиция) L1 в гене аденоматозного полипа (adenomatous polyposis coli, АРС), что лежит в основе формирования тубулярных аденом толстой кишки и колоректального рака. В результате такой транспозиции выключается функция данного белка и наступает постоянная активация сигнального молекулярного пути, приводящего к пролиферации клетки и развитию новообразований (cм. рис. 5).
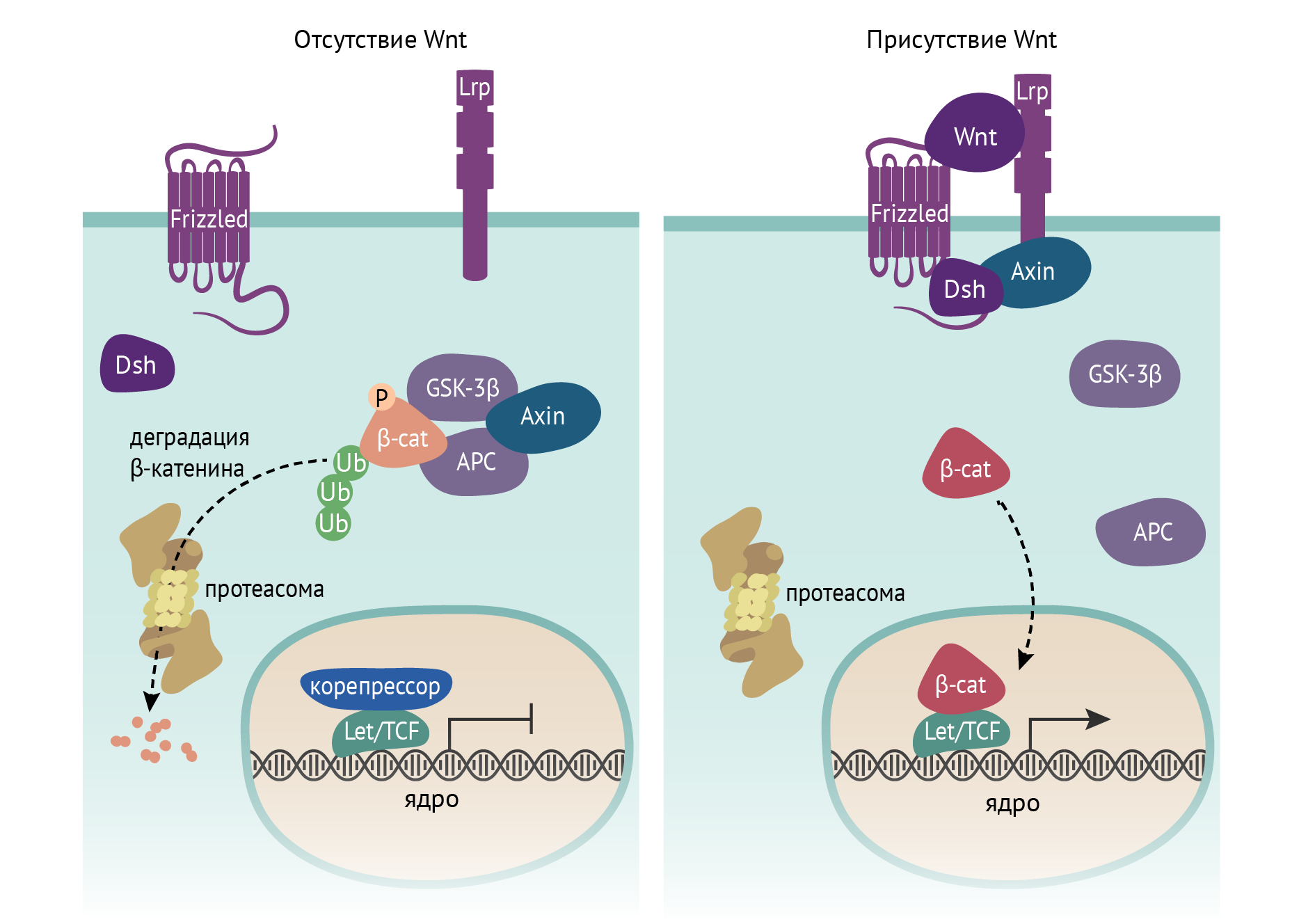
Рисунок 5 | Транспозоны. Выпадение функции белка АРС в результате транспозиции L1 с затрагиванием гена АРС приводит к конститутивной (постоянной) активности β-катенинового сигнального пути и развитию аденом и карцином толстой кишки. Белок АРС является ключевым компонентом мультибелкового комплекса, ингибирующего β-катенин путем запуска его протеасомной деградации.
Как уже было сказано, транспозоны имеют важное значение в онтогенезе, и их роль в развитии заболеваний является лишь обратной стороной медали нашего биологического разнообразия. В некоторых статьях раскрывается значение транспозонов в эволюции и онтогенезе нейронов [20]. Без транспозонов невозможна реаранжировка генома лимфоцитов, необходимая для обеспечения разнообразия синтеза антител и рецепторов к различным антигенам [1, 3, 6]. Когда-нибудь выйдет отдельный лонгрид на эту тему, но сейчас нам нужно двигаться дальше.
Повторы простых последовательностей (simple sequence repeats, SSR)
В геноме можно найти малопонятные по своему значению элементы, и одними из таких являются SSR. В структурном отношении это короткие последовательности пар нуклеотидов (приблизительно штук 10), которые повторяются миллионы раз. Они ничего не кодируют, но играют сателлитную (вспомогательную) роль, определяя ряд очень важных функций в клетке. В частности, такие сателлитные последовательности локализованы в теломерах и в центромере хромосом, что позволяет реализовывать процессы, происходящие при делении клеток [3].
Говоря о структуре хромосом, нельзя пройти мимо центромер.
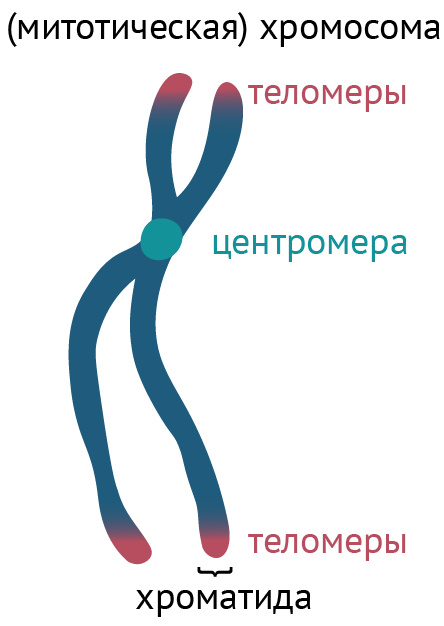
Рисунок 6 | Упрощенное изображение хромосомы
Центромера — это участок хромосомы, который соединяет собой сестринские хроматиды (см. рис. 6), но главное даже не это. Центромера является местом прикрепления белков, связывающих хромосомы и веретено клеточного деления, тем самым обеспечивая расхождение хроматид во время деления клеток. Это необходимо для распределения генетического материала по дочерним клеткам.
Сателлитные последовательности имеют важное клиническое значение. Опухолевый процесс в ряде случаев сопровождается возникновением так называемой микросателлитной нестабильности, когда в сателлитных участках возникает бесчисленное множество мутаций. Такое бывает при наличии дефектов в системе репарации, и данный феномен является предиктором высокой иммуногенности опухоли (продукции опухолью множества антигенов, способных индуцировать иммунный ответ), а также эффективности иммунотерапии — специфического варианта лекарственного лечения опухолевых заболеваний, который реализуется путем ингибирования контрольных точек иммунного ответа и помогает клеткам иммунной системы (Т-киллерам, натуральным киллерам, макрофагам) уничтожать опухолевые клетки. Положительная предиктивная роль микросателлитной нестабильности в отношении эффекта иммунотерапии продемонстрирована во множестве исследований и обзоров [15].
Оставляю тебя с этой информацией, и как только ты вернешься, мы продолжим постигать азы биохимии.
Белки хроматина
Топоизомеразы
ДНК в ходе репликации может подвергаться сверхспирализации. Этот вариант сворачивания ДНК не позволяет считать с нее что-либо. В состоянии сверхспирализации молекула ДНК наиболее стабильна в энергетическом отношении. То есть она приобретает такую конформацию, которая требует минимального (насколько это возможно) количества энергии. Это не ее причуда: данный феномен имеет термодинамическое обоснование.
Чтобы считать с нее информацию или продолжить репликацию, эту суперспираль нужно распаковать, причем это одинаково важно как в прокариотических (бактериях), так и в эукариотических клетках. Для этого есть ферменты, получившие название топоизомераз (рис. 7), которые различаются у прокариот и эукариот. Без топоизомераз клетка не может экспрессировать гены, что приводит к ее смерти [3].
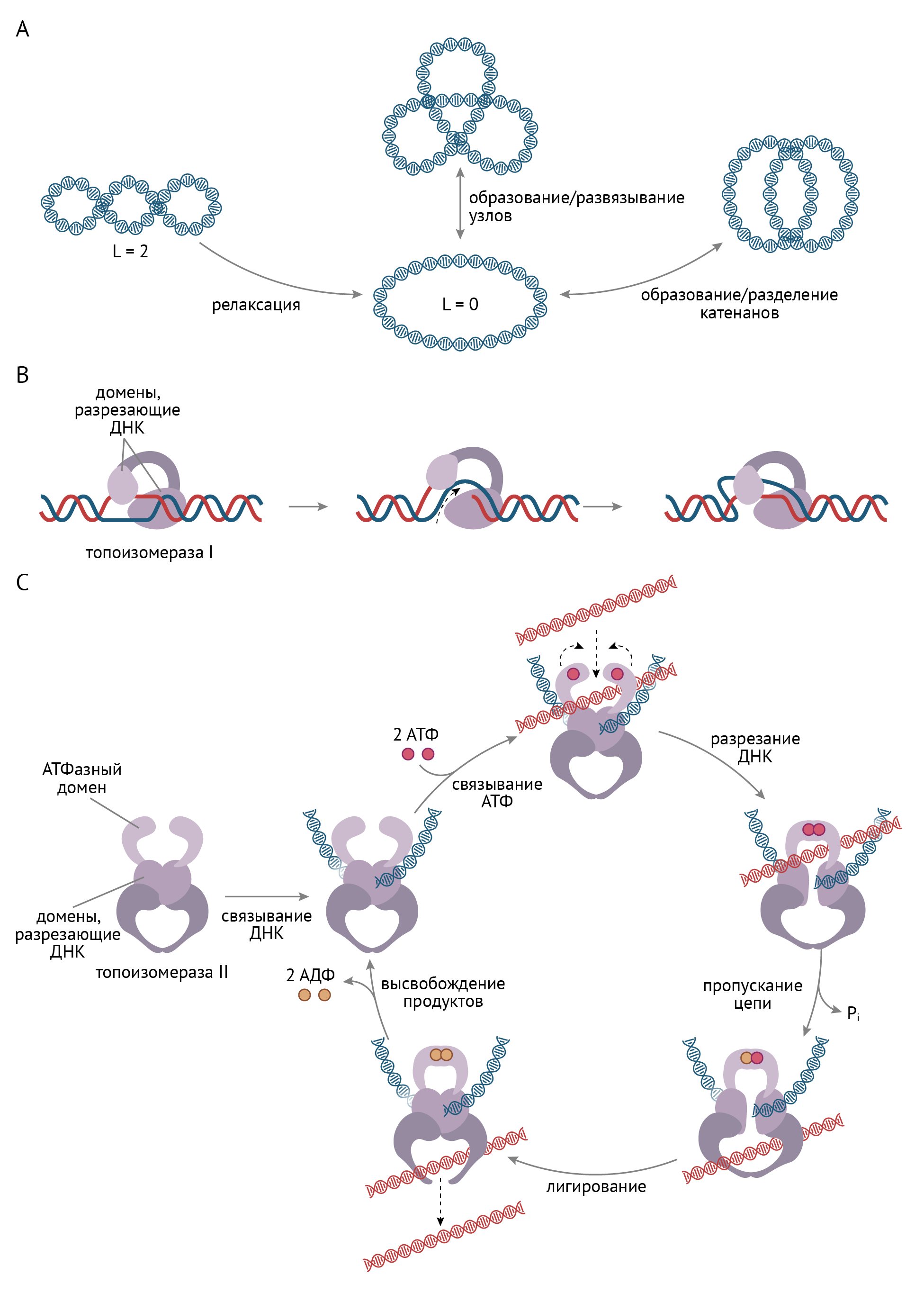
Рисунок 7 | Сверхспирализация ДНК в ходе репликации и функция топоизомераз
Как ясно из рис. 7, топоизомеразы нужны для того, чтобы молекула ДНК не превратилась в непонятную кашу, информацию с которой невозможно считать. К концу статьи ты также узнаешь, как и почему топоизомераза стала мишенью для ряда очень важных фармакологических препаратов.
Гистоны и современные представления о пространственной организации хроматина в клетке
Гистоны выводят ДНК на новый уровень, накручивая ее на себя самым причудливым образом. В этом участвует несколько фракций гистоновых белков (Н1, Н2А, Н2В, Н3, Н4). Гистоны глубоко консервативны, то есть их структура идентична гистонам более простых организмов, что для меня (убедительный уровень аргументации, правда?) подчеркивает высокое биологическое значение этих белков [1, 3, 8]. Однако как именно гистоны укладывают ДНК в более компактную структуру, которая не мешает клетке выполнять ее функции, не вполне ясно и сейчас.
Что касается третичной структуры ДНК, на сегодняшний день наши представления довольно размыты. Во многих учебниках биологии и биохимии (преимущественно русскоязычных) отражена иерархическая модель укладки ДНК. Согласно этой модели, ДНК-нуклеосомы размером 11 нанометров складываются в 30 нанометров, а затем в волокна размером от 120 до 700 нм.
Проблема в том, что данная модель была продемонстрирована в условиях in vitro. То есть из клетки были удалены другие компоненты, применялись детергенты и растворы с очень высоким содержанием солей. Поэтому вопрос, как же на самом деле уложен хроматин в клетке, долгое время оставался открытым.
Пролить свет на этот вопрос получилось у группы исследователей с помощью особого метода визуализации — ChromEMT [14]. Исследователи применили осмиофильный полимер, который позволил избирательно увеличить контрастность хроматина и лучше рассмотреть его пространственную организацию при помощи электронного микроскопа.
С помощью этой методики исследователям удалось выяснить, что диаметр хроматина в разных фазах клеточного цикла колеблется от 5 до 24 нм и что сам хроматин представляет собой неупорядоченную цепь с различными диаметрами (от 5 до 24 нм) с множеством вариантов расположений нуклеосомных частиц, бесчисленным количеством структурных конформаций и способностью изгибаться в различных плоскостях для достижения компактной ориентации в ядре. Для того, чтобы укомплектовать хроматин в ядре, не нужно формировать из него дискретные волокна более высокого порядка (в 30, 300 нм и т. д.), как считалось ранее. Нуклеосомные частицы диаметром 30 и более нм в различных фазах клеточного цикла найти просто не удалось [13]. Ниже прикреплена иллюстрация, изображающая трехмерную структуру генома в клеточном ядре (рис. 8, 9).
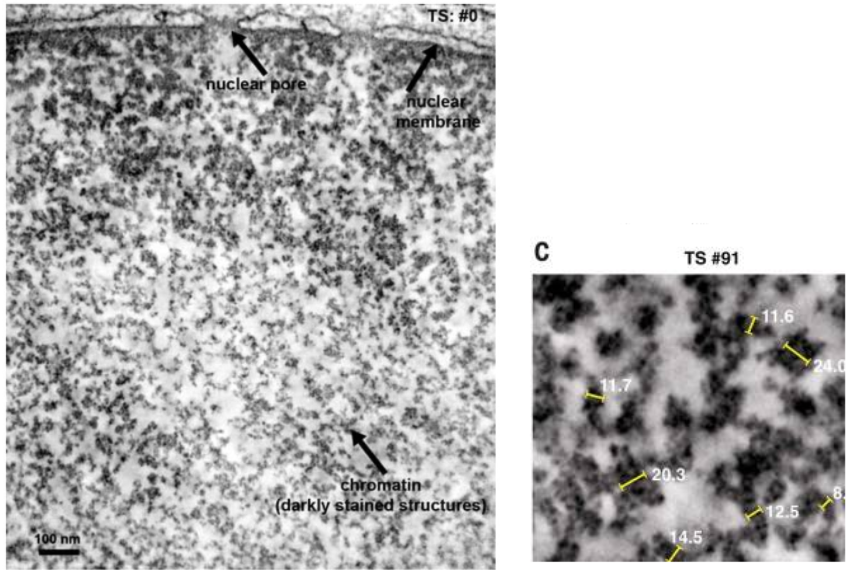
Рисунок 8 и 9 | Укладка хроматина в ядре. Данные полученные методом визуализации ChromEMT.
В целом функция гистонов сводится к компактному упаковыванию ДНК, а также к регуляции процессов считывания ДНК: репликации и транскрипции.
Структура гистонов изменяется при действии на них различных транскрипционных факторов. Эти факторы химически модифицируют N-конец гистона (путем ацетилирования, фосфорилирования, деацетилирования, метилирования), что ведет к изменению конформации гистоновых белков в целом. В ряде случаев это приводит к увеличению доступности соответствующих генов для процессов транскрипции. В виде упрощенной схемы это выглядит так (рис. 10):
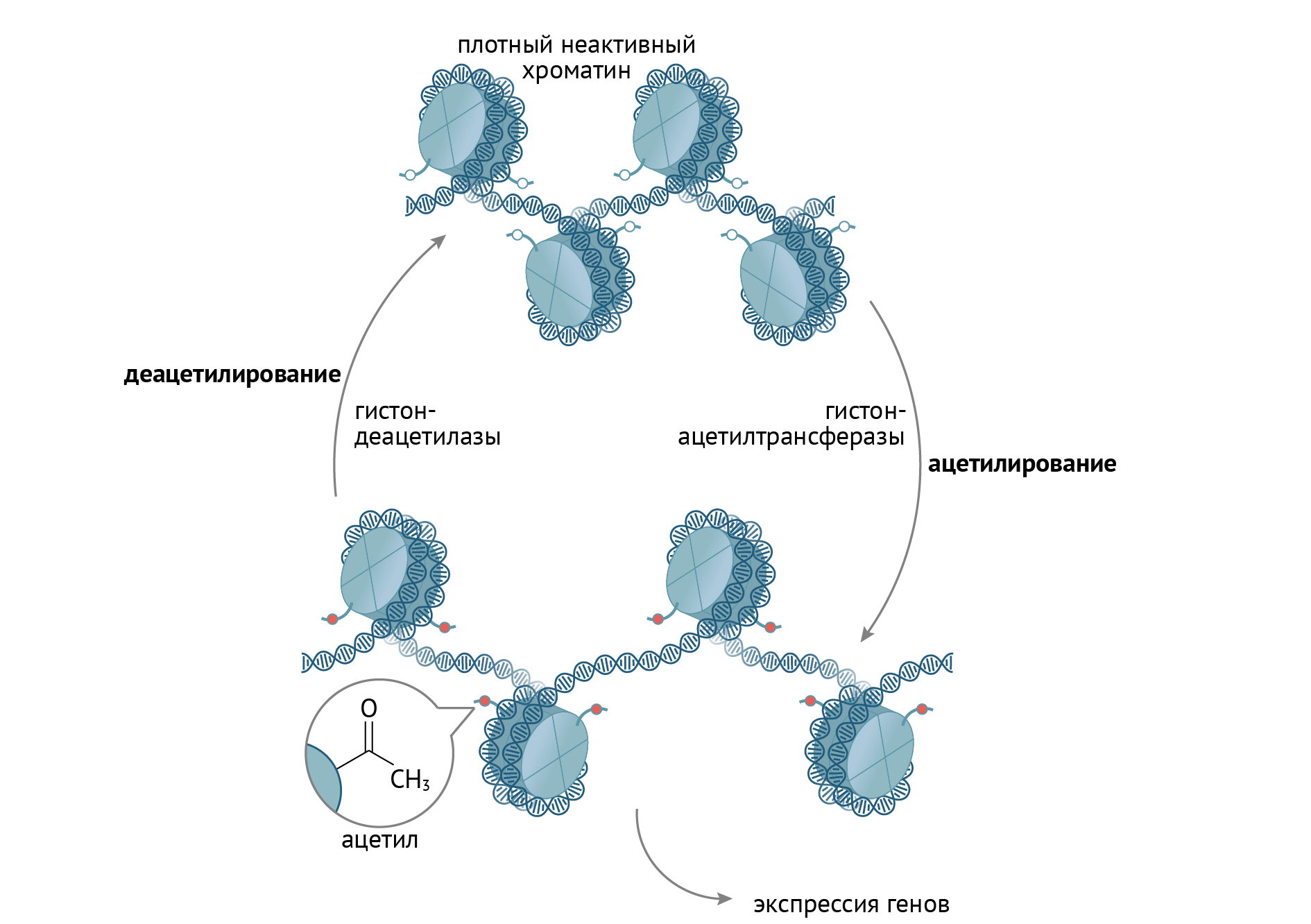
Рисунок 10 | Химическая модификация гистонов, лежащая в основе эпигенетической регуляции транскрипции генов
Метаболизм ДНК. Репликация
Мы подошли к самому сложному и интересному. Проследим цепь событий. У нас есть клетка, которая живет своей жизнью и которой, в общем-то, все равно, что происходит вокруг нее: она выполняет свою функцию, перебрасываясь с другими клетками цитокинами и метаболитами, и меряется с ними размерами своих цитоподий. Внезапно она получает «пинок» — молекулярный сигнал от старшего товарища по ткани в виде паракринных факторов роста (например, EGF — эпидермальный фактор роста; IGF-1 — инсулиноподобный фактор роста 1 и др.) или ловит своим рецептором гормон (например, эстрадиол). При воздействии на клетку вышеуказанных молекул она понимает только одно: ей приказано делиться. Химический сигнал, словно бестактное восклицание соседки: «А когда замуж?», выводит клетку из состояния покоя, когда воздействует на ее рецепторы к факторам роста. Серия реакций внутри клетки приводит к тому, что гистоны (после ацетилирования) всеми фракциями отсоединяются от ДНК, делая ее доступной для репликации, и, неожиданно, словно твое отчисление из вуза, наступают пресинтетическая и синтетическая фазы клеточного цикла. Теперь перед нами — обнаженная ДНК в виде двойной спирали, из которой нужно сделать таких две. На первый взгляд миссия более чем невыполнимая, но, как и везде, нужно подходить к этому процессу последовательно. Предлагаю не тянуть с этим.
В главных ролях:
- ДНК-полимераза;
- ДНК-хеликаза;
- ДНК-топоизомеразы;
- ДНК-праймаза;
- ДНК-гираза;
- ДНК-связывающие белки.
Во второстепенных ролях:
- Факторы, инициирующие транскрипцию ДНК (их великое множество);
- Белки клеточного цикла (о которых мы поговорим позднее).
Этапы репликации ДНК:
- инициация;
- элонгация;
- терминация.
Инициация
Уже известный нам белок, ДНК-хеликаза, присоединяется к точке инициации на молекуле ДНК. Эта точка характеризуется множеством ТА-пар в структуре и является сайтом связывания репликативного комплекса. Это связывание возможно после предварительной денатурации данного участка ДНК. Далее хеликаза движется от 5' к 3'-концу, раскручивая ДНК по мере продвижения. Параллельно действуют не только ДНК-связывающие белки, удерживающие разъединяющиеся цепи ДНК от необдуманных поступков, но и уже известные топоизомеразы (ДНК-гираза или ДНК-топоизомераза 2), которые снимают топологическое напряжение (стремление к спирализации и замыканию в себе) с цепей ДНК (рис. 11) [1, 3, 6, 8].
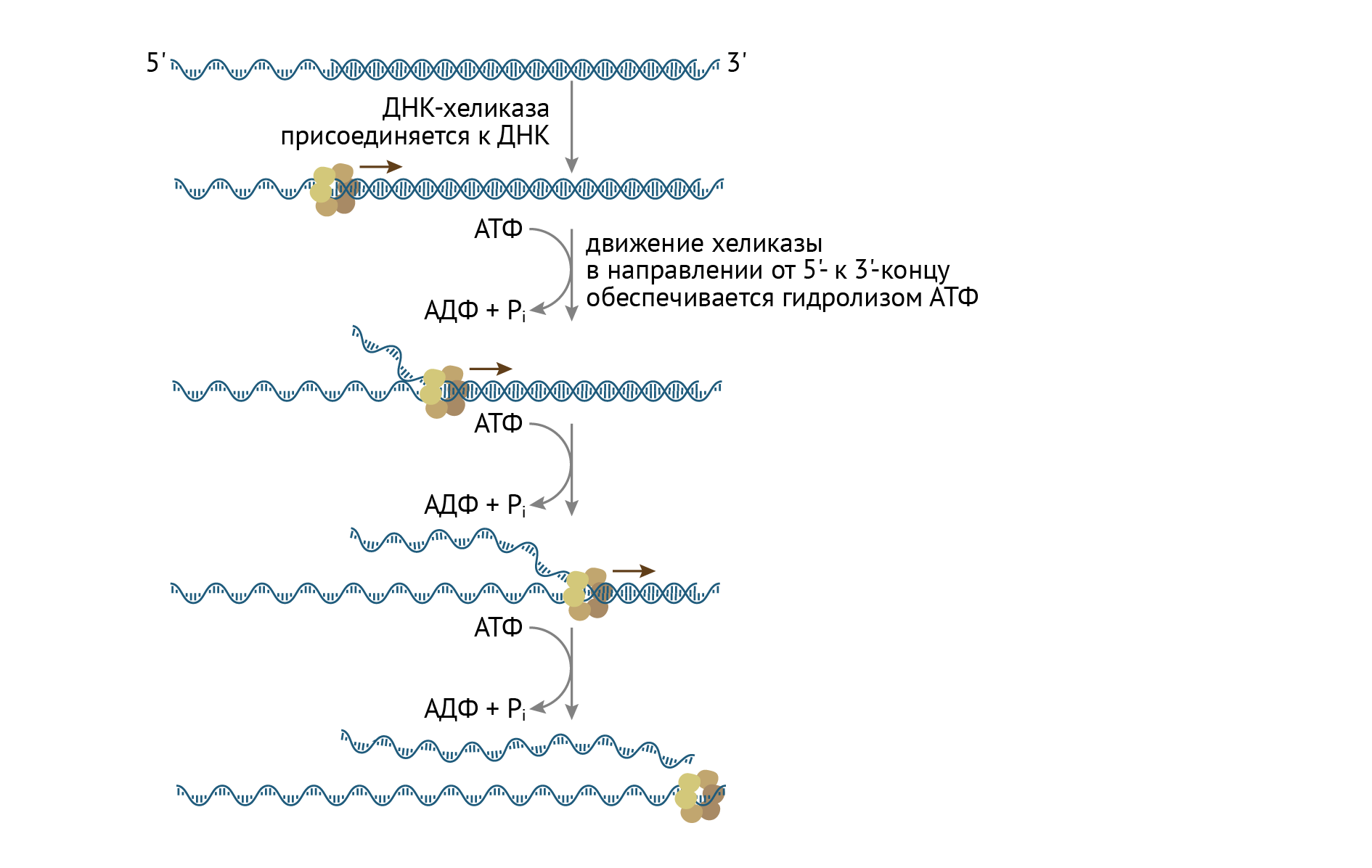
Рисунок 11 | Процесс расплетания двойной спирали ДНК с помощью ДНК-хеликаз и ДНК-связывающих белков
ДНК-полимераза не спешит работать. Она до последнего будет тянуть с синтезом нуклеиновой кислоты. Связано ли это с ее тенденцией к созидательному созерцанию или с ее природным высокомерием, но она возьмется за работу только после того, как будет установлен праймер. Праймер — это своеобразная РНК-затравка, фрагмент РНК, синтезированный РНК-праймазами. Он нужен для того, чтобы ДНК-полимеразе было что удлинять, ибо главная причина ее долгого включения в работу — неспособность синтезировать РНК-фрагмент de novo, то есть с нуля. Она может лишь удлинять имеющийся фрагмент. С формированием праймеров связан переход фазы инициации в фазу элонгации (удлинения) цепи.
Элонгация
ДНК-хеликаза связывается с РНК-праймазой, и это взаимодействие обусловливает синтез лидирующей цепи ДНК (рис. 12) [3].
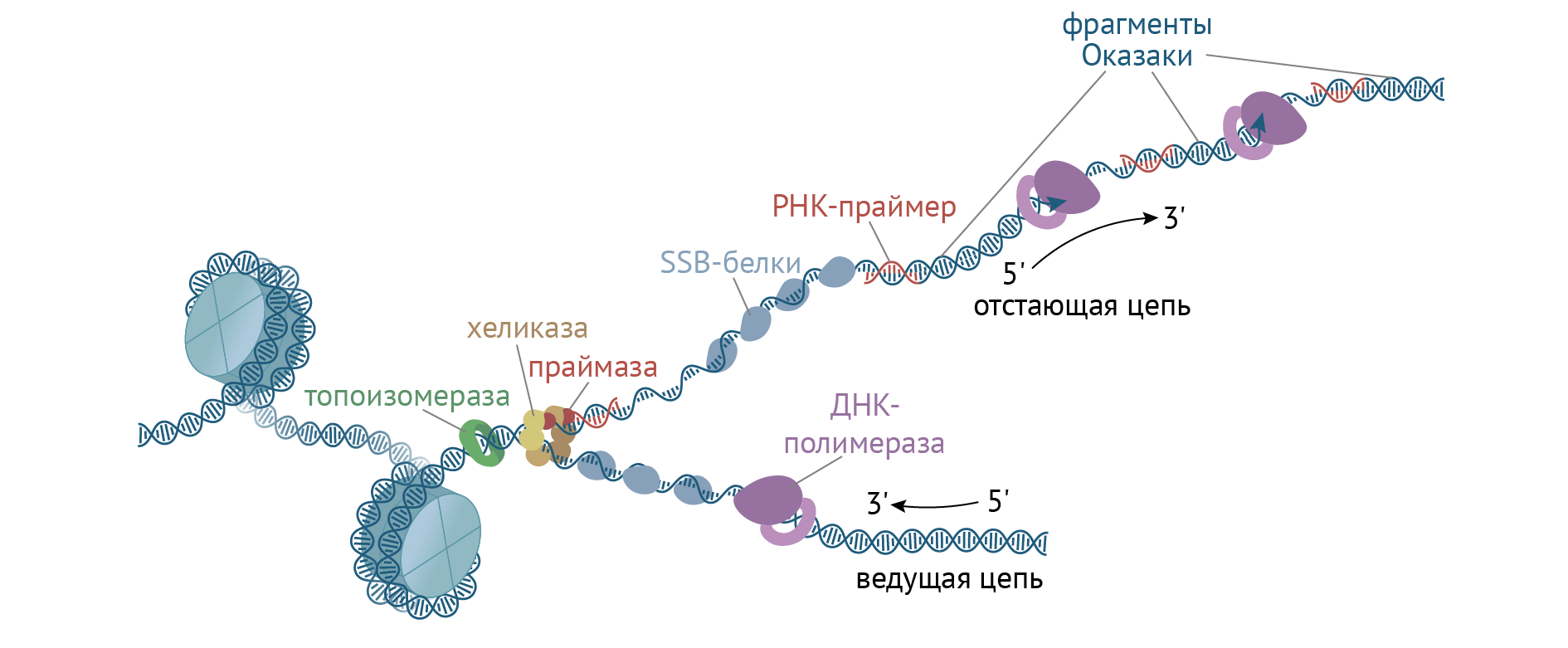
Рисунок 12 | Фаза элонгации
Синтез лидирующей цепи осуществляет ДНК-полимераза 3. Синтез на данной цепи осуществляется непрерывно, путем простого присоединения дезоксирибонуклеотидов к 3'-концу предыдущего нуклеотида. Это называется синтезом от 5' к 3'-концу.
Но события разворачиваются так, что ДНК-полимераза 3 работает на два фронта и синтезирует также и отстающую цепь. ДНК-полимераза способна работать только в направлении от 5' к 3'-концу. Отстающая цепь характеризуется противоположным направлением нуклеотидов, и, если бы ее синтез тоже был непрерывным, репликативная вилка бы полностью распалась. А вот РНК-праймаза может синтезировать цепи в направлении 3'-5'. Она делает небольшой фрагмент, который затем просто удлиняется ДНК-полимеразой. Такие кусочки называются фрагментами Оказаки. РНК-участок потом вырезается эндонуклеазой и РНК-азой. Но что самое шокирующее: синтез происходит одновременно и скоординированно. А ты жалуешься, что тебе трудно совмещать учебу с работой!
Секрет успешной работы ДНК-полимеразы 3 прост: на отстающей цепи также работает комплекс ДНК-хеликазы и ДНК-праймазы. Все вместе это можно назвать праймосомой. Далее фрагменты Оказаки сшиваются ДНК-лигазой [1, 3].
Получилось много текста, поэтому резюмирую:
- Для начала репликации нужно найти сайт связывания репликативного комплекса.
- ДНК-хеликаза необходима для разъединения цепей, иначе считать их невозможно.
- ДНК-праймаза и ДНК-полимераза необходимы для синтеза и удлинения нуклеиновой кислоты (ДНК), а ДНК-топоизомераза и ДНК-связывающие белки — для того, чтобы цепи ДНК не приняли «нечитаемую» конформацию.
- Одна дочерняя цепь синтезируется непрерывно, другая — прерывисто с помощью фрагментов Оказаки.
Нюансы и интересные факты
* Теломеры и теломеразы. В ДНК есть также концевые фрагменты, состоящие из множества повторяющихся пар нуклеотидов и не несущие существенной смысловой нагрузки. Эти фрагменты несут название теломеров (см. рис. 6).
В ходе репликации эти концевые теломеры не воспроизводятся полностью, и с каждым делением клетки теломеры постепенно укорачиваются на 5–6 пар нуклеотидов. В контексте данного вопроса выделяют такое понятие, как лимит Хейфлика — число делений, при которых еще есть чему укорачиваться. В среднем после 50–52 делений клетки теломеры укорачиваются до такой степени, что репликация в определенный момент становится невозможной, и клетка останавливается в G0-фазе (фаза покоя) клеточного цикла или запускает программу апоптотической гибели. Выходит, что по достижении данного лимита становится невозможной регенерация клеток и тканей организма. Считается, что это один из ведущих факторов, обусловливающих старение организма и конечность нашего существования (об этом говорит эпигенетическая теория старения). В этом есть свой биологический смысл, ведь при большом количестве делений происходит накопление структурных нарушений в ДНК — мутаций, которые способны стать причиной развития опухолевых заболеваний.
Однако существуют клетки (например, эмбриональные, половые и опухолевые), в которых есть теломераза — фермент, который достраивает теломеры в ДНК [2, 3]. Такое достраивание теломер делает опухолевые клетки бессмертными, поскольку возможное количество делений клеток при наличии теломеразы становится практически неограниченным.
* Топоизомеразы и мишени для фармакологических препаратов. В бактериальных клетках, как и в эукариотических, после репликации хромосомы принимают конформацию, которая наиболее выгодна в термодинамическом отношении (требует минимального количества энергии для ее поддержания), но не выгодна в отношении жизнедеятельности клетки. Что это значит? По окончании репликации цепи ДНК переплетены между собой таким образом, что транскрипция (образование мРНК в результате считывания генетической информации с ДНК) невозможна: ферментам, обеспечивающим процесс транскрипции, просто не подобраться к необходимому участку ДНК, поскольку она упакована для этого самым неподходящим образом, в виде конгломерата. Этот конгломерат цепей не распутать без участия ДНК-топоизомеразы IV (топоизомеразы II типа). Они вносят кратковременный разрыв в обе цепи ДНК, и одна из цепей ДНК может пройти через этот разрыв (рис. 7) [3].
* Топоизомераза IV является важной лекарственной мишенью для противомикробных препаратов широкого (очень) спектра действия — хинолонов и фторхинолонов. Препараты избирательны по отношению к топоизомеразе IV и не проявляют интереса к топоизомеразам эукариот, что делает их применение очень удобным (и подчас спасает жизни). Как ты понимаешь, если вновь образовавшиеся хромосомы похожи на сыр «косичка», адекватно считать с них информацию бактериальная клетка не может. И ей ничего не остается, как погибнуть. Несмотря на селективность данных препаратов, их нельзя назвать абсолютно безопасными, но об этом ты узнаешь чуть позже на фармакологии [3, 9].
Синтез РНК и полипептидов
Синтез РНК. Транскрипция
Синтез РНК в общих чертах напоминает репликацию ДНК с некоторыми нюансами:
- РНК-полимераза самодостаточна: ей не нужен праймер для начала работы.
- Субстратом для образования РНК являются рибонуклеотиды с аденином, гуанином, цитозином и урацилом (вместо тимина).
- Если при репликации копируется вся цепь ДНК, то в случае с транскрипцией читается лишь определенный участок ДНК, а именно ген, кодирующий определенный полипептид.
- Синтез РНК начинается после связывания РНК-полимеразного комплекса с промотором. Это участок ДНК, который сигнализирует: синтез данного полипептида нужно начать отсюда. Промотор богат ТАТА-последовательностями нуклеотидных пар, за что получил кличку «ТАТА-бокс» [1, 3].
Даже стадии транскрипции такие же, как и стадии репликации: инициация, элонгация и терминация. Но опять же с некоторыми нюансами.
Инициация
Сначала нужно образовать транскрипционный комплекс. В него входят:
- белки, обладающие хеликазной (т. е. «распутывающей нити») активностью;
- белки, связывающие ТАТА-бокс;
- белки, стабилизирующие всю эту машину;
- белки, связывающие РНК-полимеразу II с ДНК в области ее промотора;
- РНК-полимераза II типа (да, у эукариот их три) и белки, которые ее фосфорилируют.
Более того, в этой машине для синтеза РНК есть даже белки, которые обладают репаративной активностью, т. е. во время транскрипции осуществляют эксцизионную репарацию. Эксцизионная репарация — метод устранения структурных нарушений в ДНК (мутаций) путем «вырезания» или так называемой эксцизии участка ДНК, имеющего структурный дефект, и последующей достройки недостающего участка по принципу комплементарности.
В целом все эти белки можно обозвать факторами транскрипции, которым на Западе дали различные буквенные обозначения, но они сейчас нас мало интересуют. Получается здоровая такая бюрократическая машина, но с совсем не бюрократическими целями [1].
Инициация запускается в области промотора, но сама структура промотора не имеет ничего общего с кодирующей областью ДНК и с кодирующим полипептидом соответственно. Запускается она путем множественного фосфорилирования РНК-полимеразы двумя участниками транскрипционного тандема.
Элонгация
Удлинение цепи без регистрации и СМС путем простого присоединения нуклеозидтрифосфатов по направлению к 3'-концу. РНК-полимераза II типа на стадии элонгации обладает нереально высокой активностью, которая поддерживается связанными с нею белками (факторами элонгации) [3].
Терминация
Когда весь здоровенный комплекс натыкается на сайты терминации, РНК-полимераза дефосфорилируется и слезает с ДНК, выдавая готовую прематричную РНК. Из нее уже можно получить что-то дельное [8].
Комментарий
У эукариот выделяют три РНК-полимеразы, но главную роль в генной экспрессии действительно играет именно РНК-полимераза II. РНК-полимераза II образует прерибосомные РНК, которые участвуют в образовании рибосом — подпольных фабрик по синтезу белка. РНК-полимераза III участвует в образовании специализированных РНК, функции которых до конца не охарактеризованы [3].
Нюансы и интересные факты
* РНК-полимеразы тоже плачут. Нечасто используемый, но чрезвычайно сильный антибиотик рифампицин (из группы ансамицинов) способен подавлять синтез бактериальных РНК путем связывания с -субъединицей бактериальных РНК-полимераз. В результате ее инактивации не происходит высвобождение промотора при транскрипции (по сути, не начинается транскрипция). Это позволяет использовать рифампицин в самых тяжелых случаях: при стафилококковом остеомиелите, эмпиеме плевры и даже различных формах туберкулеза [8, 9].
Процессинг как промежуточный этап на пути синтеза белка
После образования первичного транскрипта (первичной, или пре-мРНК) происходит ее обработка в виде процессинга. Он включает в себя:
Кэпирование (присоединение 7-метилгуанозина, связанного с 5'-концевым остатком мРНК). Функция этой метки — защита мРНК от рибонуклеаз, а еще он имеет значение для инициации трансляции.
Полиаденилирование — присоединение множества нуклеотидов с аденином на конец пре-мРНК, имеет ту же цель, что и кэпирование — защитить от рибонуклеаз ядра и цитоплазмы (защита недолгая, но какая есть). Полиаденилирование также способствует инициации трансляции: полиаденилированный конец мРНК способствуют правильной взаимной пространственной ориентации малой субъединицы рибосом и мРНК.
Сплайсинг. Это самая интересная часть, которая заключается в вырезании интронов — участков ДНК, не несущих в себе информации. Вновь синтезированная молекула РНК называется первичным транскриптом, и именно она подвергается дальнейшей модификации в виде сплайсинга. Вырезание интронов происходит под действием сплайсосом (кстати, они являются РНК по своей природе). На выходе мы получаем пептид, состоящий только из экзонов: в процессе сплайсинга интроны удаляются из первичного транскрипта, а экзоны соединяются с образованием непрерывной последовательности, которая и соответствует тому или иному функциональному полипептиду (рис. 13) [1, 3, 6, 8].
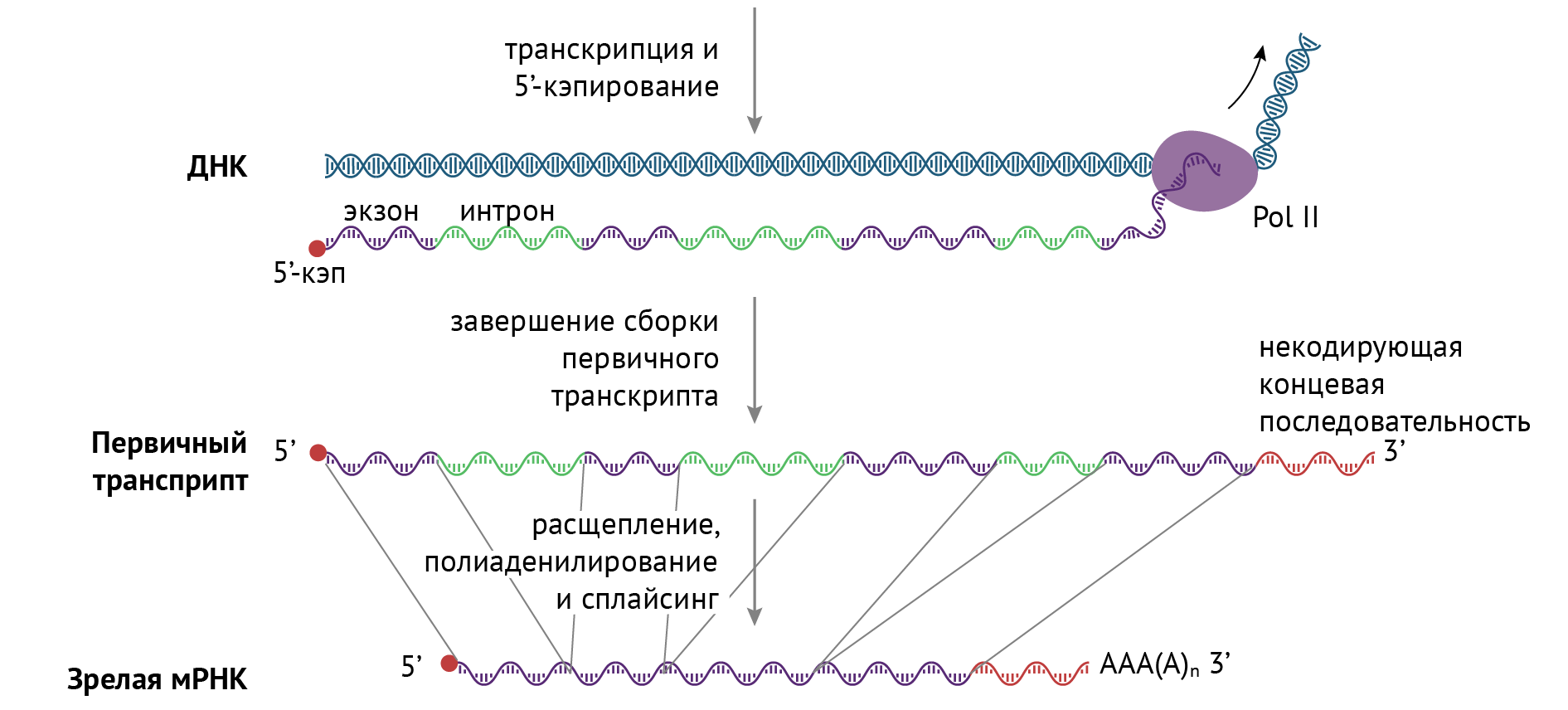
Рисунок 13 | Сплайсинг первичного транскрипта [6]
Наиболее интересно во всей этой истории явление альтернативного сплайсинга. Мы говорили о нем, когда упоминали, что клетка научилась виртуозно жонглировать экзонами и интронами и получать различные полипептиды из ограниченного числа генов. При альтернативном сплайсинге могут «нарезаться» самые разные комбинации экзонов. В итоге на выходе получаются различные белки из одного первичного транскрипта (т. е. из одного гена) [3]. Это очень удобно для клетки.
Как же это происходит? Первичный транскрипт содержит молекулярные сигналы для всех альтернативных путей сплайсинга (т. е. потенциально способен преобразоваться в любой вариант матричной РНК), но предпочтительный в данный момент путь определяется факторами процессинга — РНК-связывающими белками, которые и запускают один конкретный путь сплайсинга. Одним из наиболее ярких примеров альтернативного сплайсинга является образование антител В-лимфоцитами.
Трансляция
Этот процесс сложнее, чем кажется на первый взгляд. Для его осуществления клетке нужно сделать очень много чего.
В первую очередь нужно перевести аминокислоты в такую форму, в которой они смогут участвовать в биосинтезе белка. Для этого к аминокислоте (любой) присоединяются S-KoA группы с последующим добавлением к ней тРНК. Схематично и упрощенно процесс изображен на рис. 14 [6, 8]:
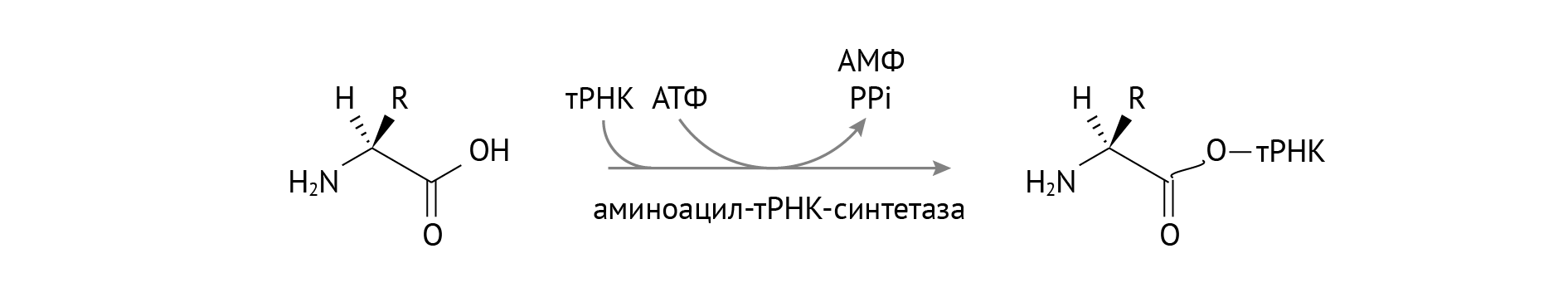
Рисунок 14 | Активация тРНК для ее участия в трансляции
Только в такой форме аминокислоты способны пройти фейсконтроль на входе в рибосомы. Для этого процесса, кстати, нужны ионы магния [1]. Без него машина под названием «аминоацил-тРНК-синтетаза» не работает. В целом весь процесс трансляции (биосинтеза белка) изображен на рис. 15.
Инициация
МРНК, образованная в ядре за счет транскрипции и процессинга, наконец-то попадает на место сходки. И сразу взаимодействует с меньшей субъединицей рибосомы и с активированной аминоацил-тРНК. После присоединения большой субъединицы рибосомы образуется инициаторный комплекс. Важно сказать, что инициаторной кислотой будет являться N-формилметионил- аминоацил-тРНК.
Но и это еще не все. Для инициации трансляции нужна энергия в виде ГТФ и присутствие факторов инициации трансляции [8].
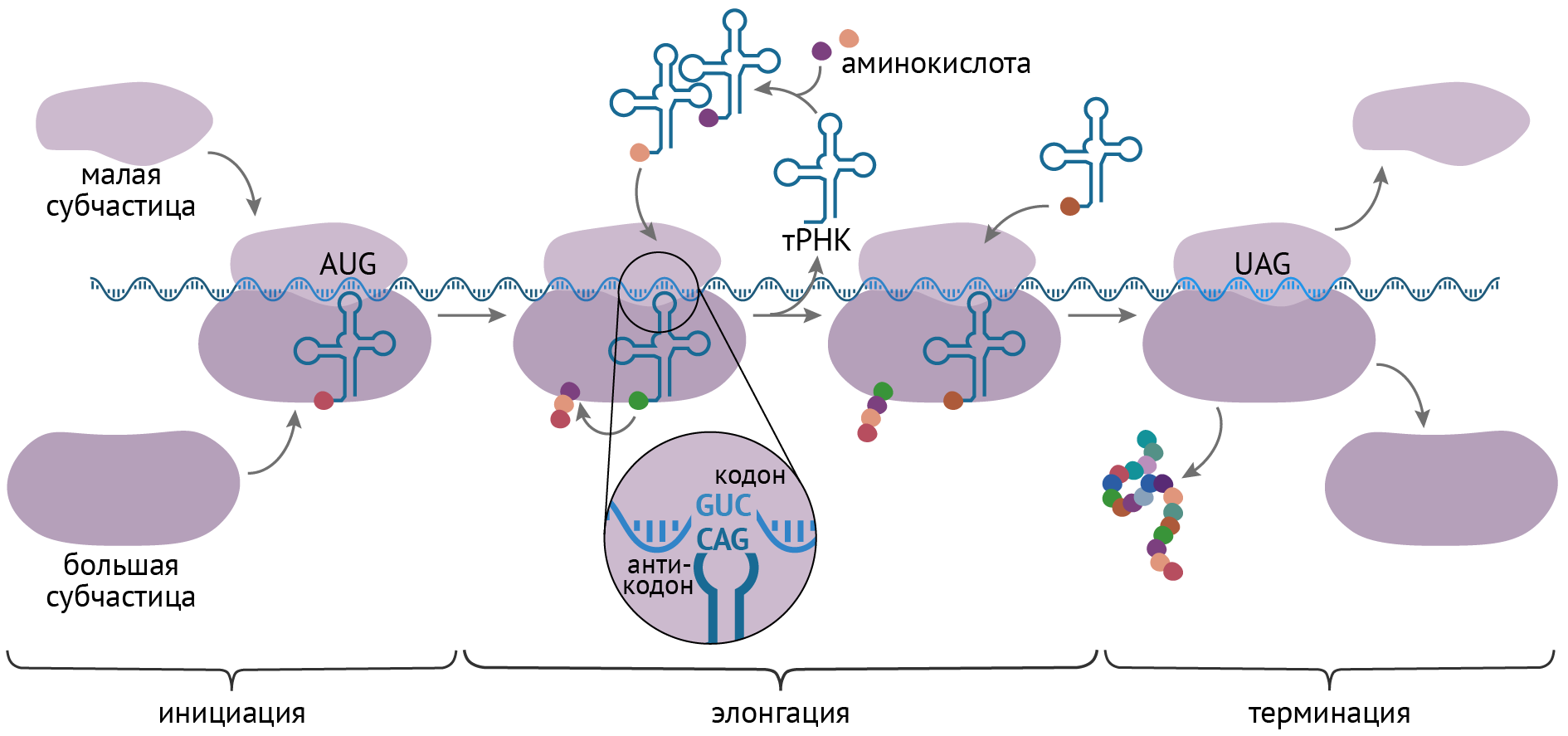
Рисунок 15 | Фазы трансляции
Элонгация
Факторы элонгации, которых в клетке достаточно, осуществляют присоединение вновь прибывающих аминоацил-тРНК к предыдущей молекуле. Здесь, кстати, тоже соблюдается принцип комплементарности (англ. complement — дополнение) между мРНК и тРНК. Примечательно, что элонгация полипептида — это процесс, который требует энергии для осуществления, но ее источником служит не АТФ, а ГТФ. Сложно сказать, с чем связаны эти гастрономические предпочтения клетки [6, 8].
Терминация
Терминация трансляции и высвобождение полипептида осуществляются тогда, когда тРНК натыкается на стоп-кодон. В результате этой встречи рибосома понимает, что никто не вечен, и распадается на субъединицы. А довольный полипептид проходит следующие круги капиталистического синтеза и попадает в ЭПР и комплекс Гольджи, где происходит укладка полипептидов и их посттрансляционный процессинг.
Укладка полипептидов. Посттрансляционный процессинг полипептидов
Для того, чтобы стать полноценными в функциональном отношении белками, полипептиды должны принять третичную (нативную) структуру. Этот этап занимает важное место в жизни белка и называется фолдингом (от англ. folding — укладка). К белкам, помогающим другому белку стать «большим», относятся белки-шапероны, или белки теплового шока.
Сворачивание белка проходит через кинетически предпочтительные стадии, определяемые особенностями первичной структуры (у разных полипептидов они разные). Это значит, что процесс будет протекать тем быстрее, чем меньше энергии необходимо. Цепочка из аминокислот не может просто так взять и превратиться в третичную структуру, это требует затрат энергии, а процесс включает определенные пространственные и химические превращения. Белковой молекуле надо помочь, и делают это белки-шапероны [3, 6]. Проследим механизм их действия по рис. 16.
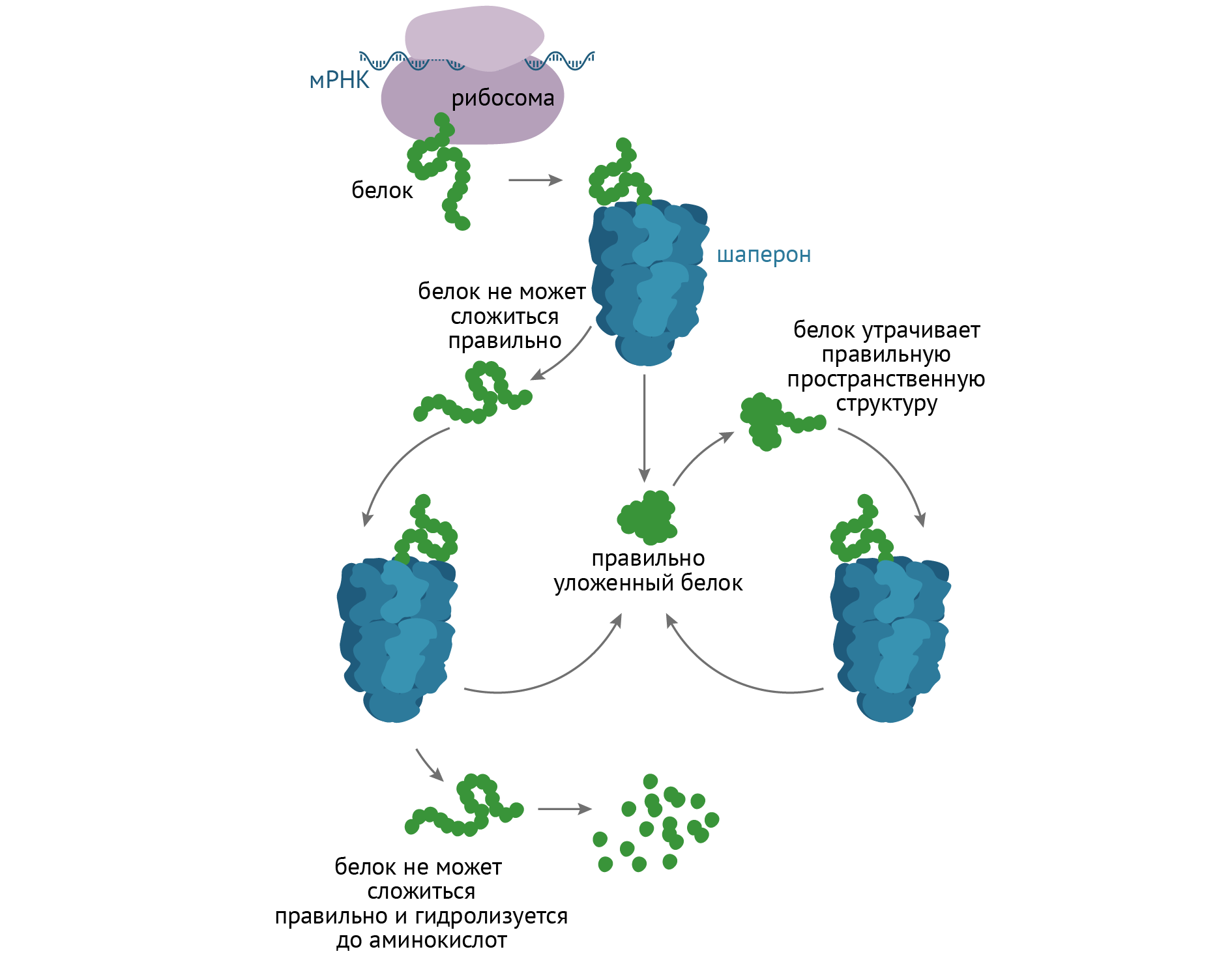
Рисунок 16 | Фолдинг белков
Шапероны предотвращают агрегацию белковых молекул путем защиты от других пептидов и связывания гидрофобных групп. Используя энергию АТФ, шапероны помогают пептиду преодолеть энергетический барьер на пути к формированию нативной белковой молекулы.
Для тех, у кого не получилось стать нормальным белком, клетка предусмотрела механизм убиквитин-протеасомной деградации [2, 6] (рис. 17).
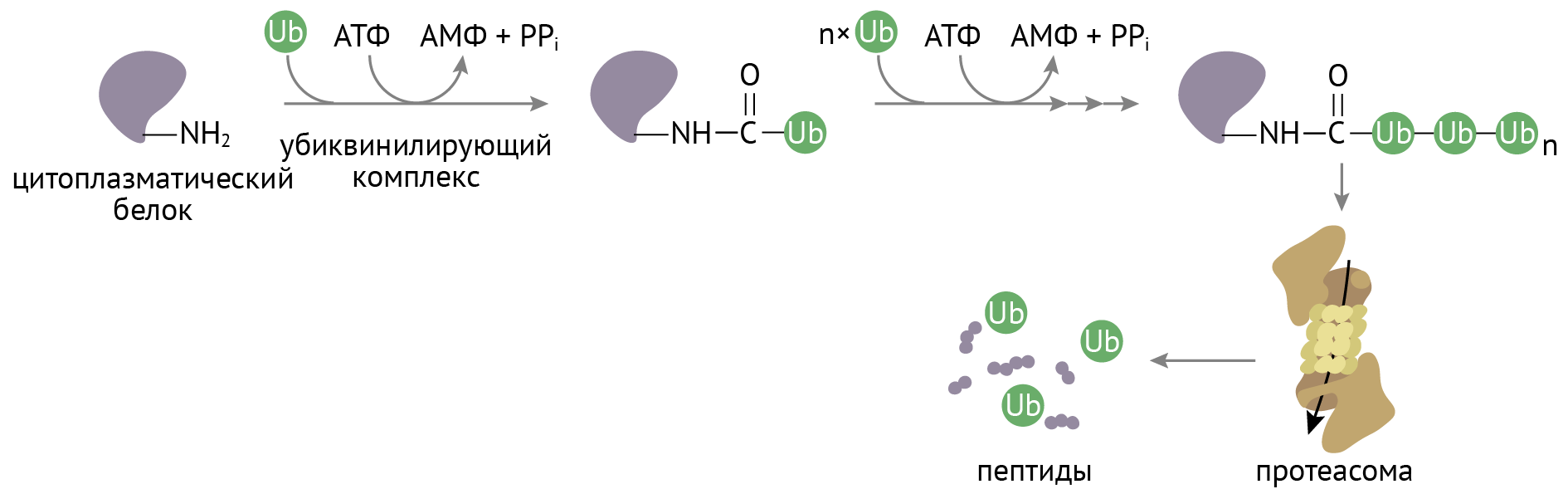
Рисунок 17 | Протеасомная деградация неудачно свернутых пептидов
К неудачному в структурно-функциональном отношении пептиду присоединяется убиквитин под действием убиквитин-активирующего фермента. С этого момента неудачный пептид является прокаженным, ведь он обладает убиквитиновой меткой. Эта метка нужна для протеасом, которые захватывают пептиды и подвергают их гидролизу. Если дефектные белки не уничтожать, то клетка заполнится хламом в виде неудачно свернутого белка (как твоя комната в общаге на второй неделе учебы), в которой невозможно будет осуществлять нормальную жизнедеятельность (ну ты понял) [2, 5, 6].
Но бывают и более благоприятные варианты развития событий. Большая часть белка подвергается другому варианту посттрансляционного процессинга в ЭПР и в комплексе Гольджи. Здесь возможны самые разные варианты химического извращения, например, присоединение глюкозы и ее производных с образованием разнообразных гликопротеидов.
Клинические аспекты биосинтеза макромолекул
Антибактериальные и синтетические противомикробные препараты, угнетающие синтез белка в бактериальных клетках
Биосинтез белка и репликация ДНК, как ты мог догадаться, протекают и в бактериальных клетках, причем достаточно интенсивно. Отдельные представители размножаются со скоростью три деления в час, и это нереально быстро. Размножение бактерий сопряжено с повреждением тканей макроорганизма в месте данного безобразия, что вызывает воспалительную реакцию и становится патологоанатомической основой большинства инфекционных заболеваний бактериальной этиологии. Данные заболевания требуют назначения антибактериальных препаратов, без вариантов. Большинство этих препаратов воздействует на метаболизм белка или нуклеиновых кислот в бактериальных клетках.
Ингибиторы топоизомеразы II типа (ДНК-гиразы IV). Роль данных ферментов ты уже знаешь, но я напомню: ДНК-топоизомераза 2 типа разрывает обе цепи молекулы ДНК и пропускает в образующуюся брешь другой участок молекулы ДНК, что предотвращает спирализацию (скручивание) молекулы ДНК (рис. 7). Есть данные, что отсутствие ДНК-гиразы приводит к пространственному разобщению цепей ДНК в ходе их спирализации.
Для бактериальных клеток фермент имеет колоссальное значение, и при его отсутствии репликация попросту не начнется. Именно поэтому человечество создало препараты, способные угнетать данные ферменты, и это хинолоны/фторхинолоны [3, 6, 9].
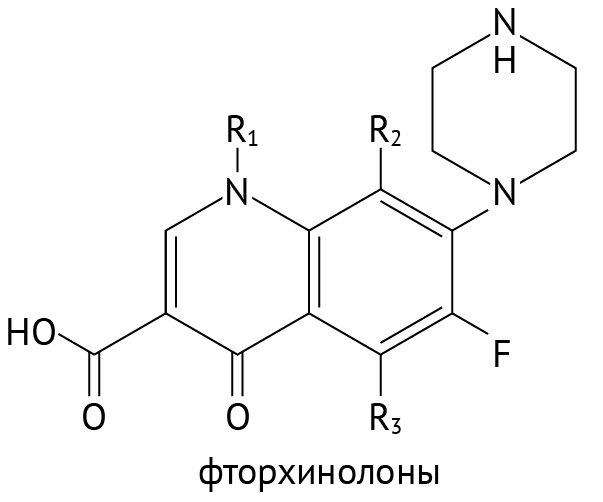
Рисунок 18 | Формула фторхинолонов
Препаратов данного класса много, наиболее «ходовые» — ципрофлоксацин, левофлоксацин, норфлоксацин, моксифлоксацин. Общим для них является достаточно широкий спектр действия, но они не действуют на анаэробные бактерии, листерии (факультативный анаэроб), спирохеты (к ним относится возбудитель сифилиса Treponema pallidum), на микобактерии (среди них — возбудители лепры и туберкулеза). Хотя стоит заметить, что офлоксацин и ципрофлоксацин проявляют определенную активность и в отношении возбудителей туберкулеза. Препараты обладают серьезными преимуществами: они хорошо проникают в ткани, даже если поступают в организм в таблетированной форме, к ним долго развивается устойчивость бактерий, у них адекватный профиль токсичности. Однако их нельзя применять у детей (только по жизненным показаниям, off-label, через консилиум), поскольку есть данные о негативном влиянии на хрящевые зоны роста [17]. У пожилых на фоне применения фторхинолонов может развиться специфическое осложнение — разрыв ахиллова сухожилия [18]. В силу широкого распространения антибиотикорезистентности фторхинолоны сейчас являются препаратами резерва и используются преимущественно тогда, когда другие антибактериальные препараты неэффективны.
Примечательно и то, что фторхинолоны не ингибируют топоизомеразы эукариотических клеток в терапевтических дозах. Это делает их сравнительно безопасными, хотя и не означает, что они лишены побочных явлений [9].
Ингибиторы биосинтеза белка бактерий. У бактерий рибосомы отличаются меньшим коэффициентом седиментации (70S vs 80S у эукариотических клеток). Существует ряд препаратов, селективно угнетающих биосинтез белка на бактериальных рибосомах.
Макролиды неравнодушны к пептидилтрансферазному центру 50S-субъединицы рибосом бактерий. Когда молекула макролида связывается с ним, нарушаются процессы транслокации аминокислот (то есть страдает фаза элонгации), и белок не собирается.
Несмотря на то, что биосинтез белка протекает во всех бактериальных клетках, макролиды не обладают достаточно широким спектром противомикробной активности. Он ограничивается преимущественно грамположительной микрофлорой. Но главным достоинством макролидов является их высокая эффективность против внутриклеточных бактерий, паразитирующих в эукариотических клетках. К таким относят микоплазмы и хламидии, в том числе уретральные.
Представители: азитромицин, кларитромицин, рокситромицин, телитромицин, старый как мир эритромицин и прочие. Препараты характеризуются удовлетворительной переносимостью и безопасностью, что обусловливает их широкое применение [3, 9].
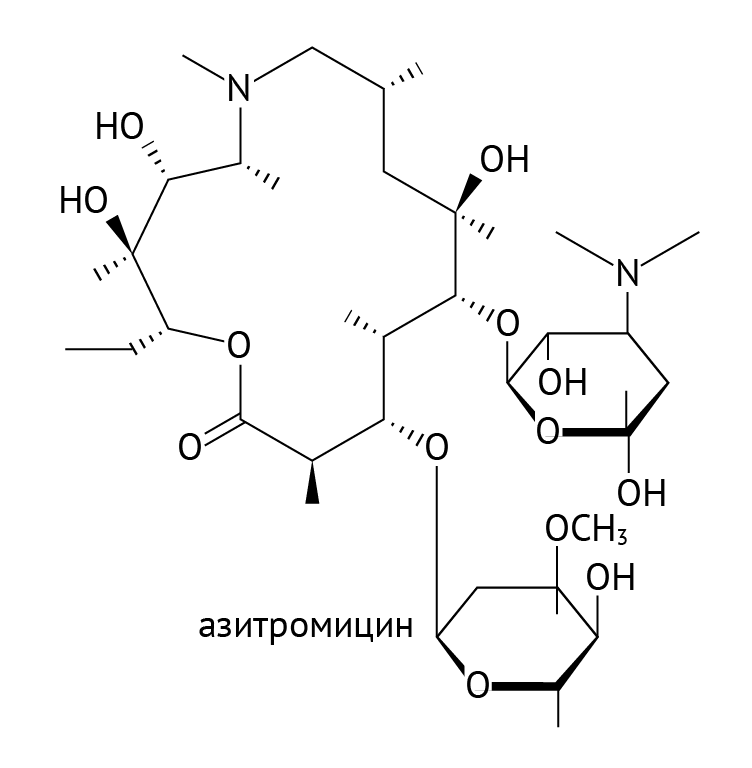
Рисунок 19 | Азитромицин
К особенностям макролидов относят их способность создавать высокую концентрацию в очаге воспаления. Макролиды внедряются в макрофаги, которые добираются до очага воспаления (под действием соответствующих хемоаттрактантов) к концу первых суток от повреждения ткани. К другой особенности относят способность макролидов проникать в клетки микоплазм и хламидий. Особо важное значение это имеет для педиатрии: макролиды — практически единственные антибиотики, которыми можно победить респираторный микоплазмоз у детей младшего возраста. Другие эффективные по отношению к микоплазме препараты у детей используются лишь по жизненным показаниям в силу их профиля токсичности.
Аминогликозиды. Угнетают 30S-субъединицу рибосом, что затрудняет движение рибосомы по нити мРНК. Также нарушают считывание кода мРНК и повреждают бактериальную мембрану клеток, но это уже другая история [9].
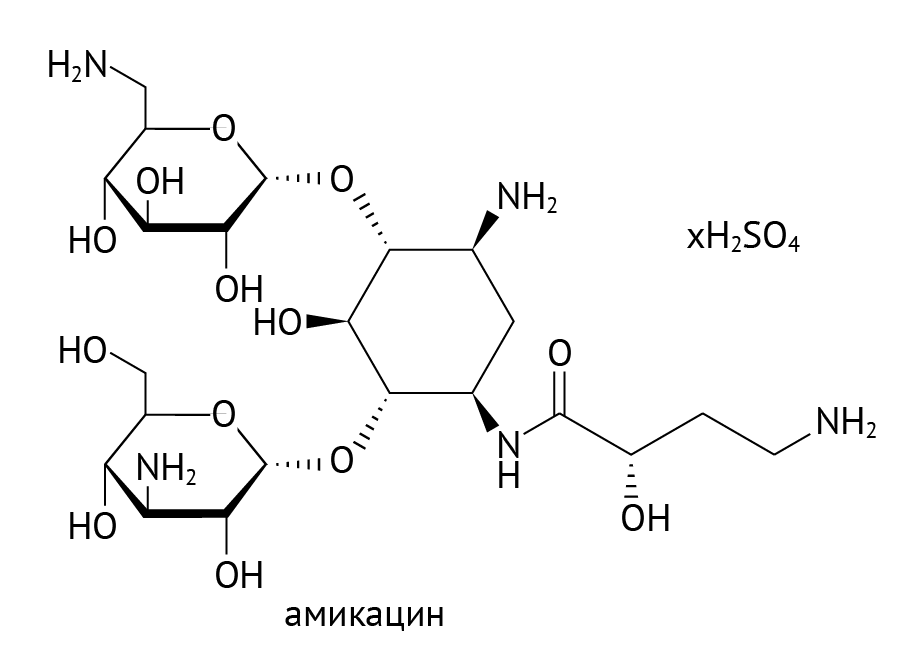
Рисунок 20 | Амикацин
Аминогликозиды применялись бы широко, если бы не их нефро-, ото- и вестибулотоксичность. Механизм повреждения слухового анализатора и вестибулярного аппарата все еще не понятен. Есть данные, что аминогликозиды активно накапливаются в пери- и эндолимфе внутреннего уха, достигают токсической концентрации в ней, угнетают биосинтез белка и нарушают целостность клеточных мембран [19]. Нефротоксичность обусловлена канальцевой секрецией аминогликозидов и токсическим повреждением эпителия канальцев почек.
Аминогликозиды — препараты с узким терапевтическим диапазоном, поэтому применяются по строгим показаниям.
Похожая ситуация сложилась с тетрациклинами. Среди них сейчас используется в основном доксициклин (рис. 21)
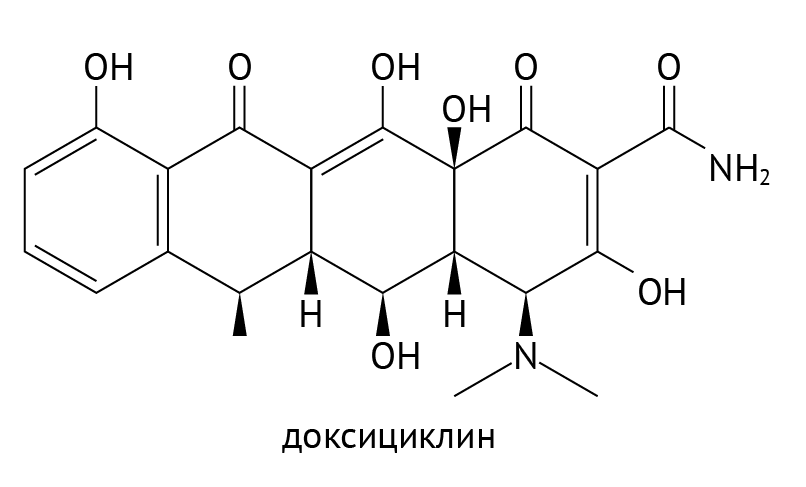
Рисунок 21 | Доксициклин
Тетрациклины обладают хорошей биодоступностью (в силу высокой липофильности) и исходно действуют против большого количества микроорганизмов. Но получилось так, что многие бактерии приобрели к ним устойчивость в результате бесконтрольного применения. Их применение у детей ограничено из-за токсического действия на хрящевые зоны роста. Препараты разрешено применять лишь после 8 лет. Тем не менее тетрациклины успешно используются в лечении некоторых опасных зоонозных инфекций, например, туляремии и бруцеллеза.
Клеточный цикл и роль структурных нарушений ДНК в онкогенезе
Рост и пролиферация клетки в обычных условиях находится под жестким контролем регуляторных систем более высокого уровня. Другими словами, пока клетке не скажут: «Делись, мать!», та не делится. Вся жизнь клетки подчинена клеточному циклу [2]. Рассмотренная нами в предыдущих параграфах репликация ДНК происходит в синтетическую фазу S, тогда как биосинтез белка может происходить во все остальные фазы. Движущей силой клеточного цикла является последовательная активация сменяющих друг друга циклинзависимых протеинкиназ, набор которых в зависимости от фазы цикла различается, во многом в результате действия на клетку различных биохимических сигналов (факторов роста, цитокинов, гормонов) [2, 3]. Схематично сигнальные каскады клетки проиллюстрированы на рис. 22.
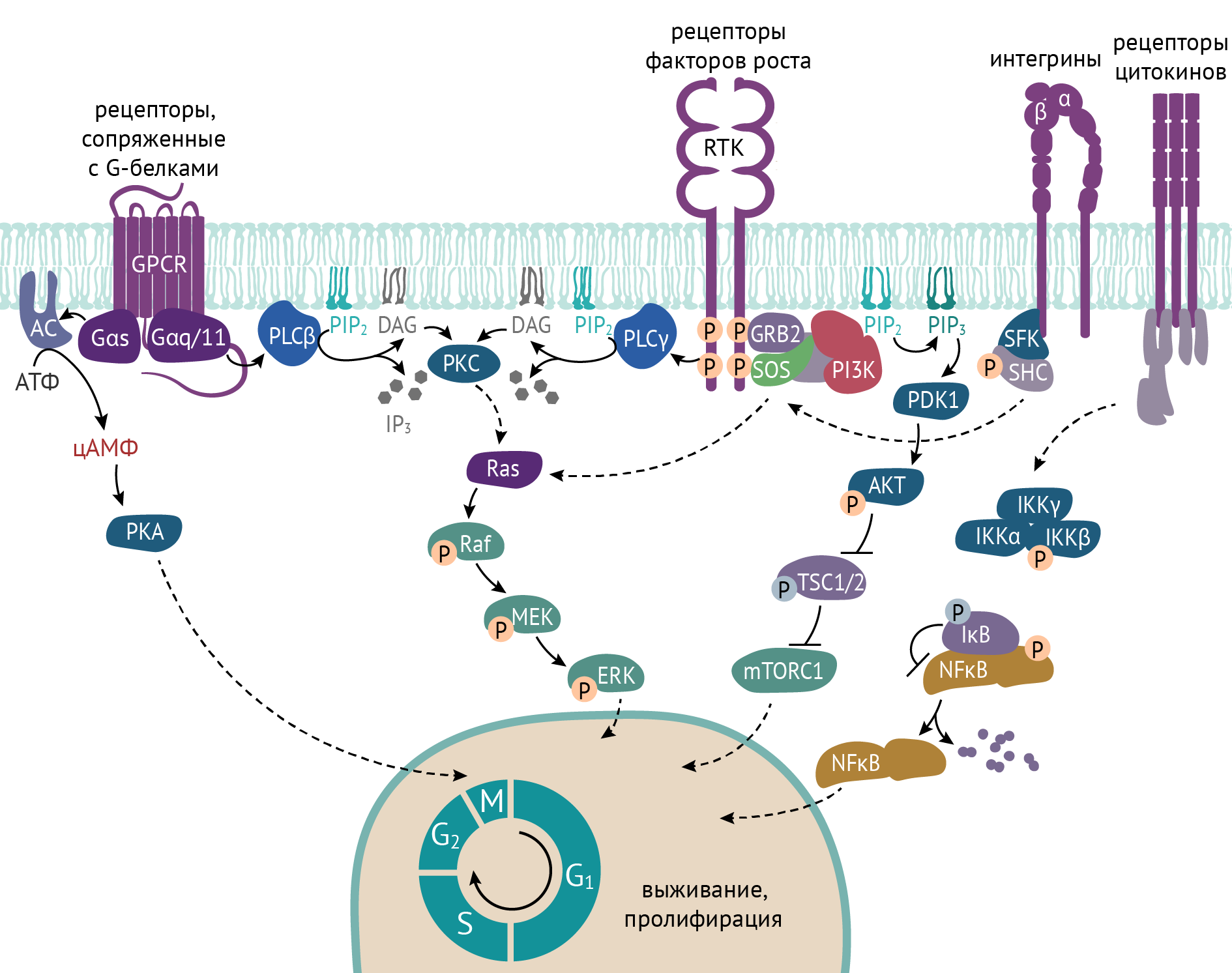
Рисунок 22 | Сигнальные каскады, запускающие деление клетки [1]
Различные рецепторы к факторам роста (эпидермальному, фибробластическому, тромбоцитарному, инсулиноподобным) при взаимодействии со своим лигандом индуцируют активацию каскада внутриклеточных биохимических реакций, приводящих к изменению транскрипции различных циклинзависимых киназ и других участников, обусловливающих смену фаз клеточного цикла [2 ,6].
Реакции, запускающие смену фаз клеточного цикла, сложны и многогранны, но их суть сводится к активации определенных транскрипционных факторов, которые в свою очередь обеспечивают считывание генов, ответственных за экспрессию белков-участников клеточного цикла [4]. В схеме ниже факторы роста обозначены как «факторы выживания».
Представляя эту схему (рис. 22), не могу не отметить, что для многих клеток взаимодействие с их микроокружением (в том числе с внеклеточным матриксом) является необходимым условием для пролиферации. К таким клеткам можно отнести эпителиальные клетки. Для инициации клеточного деления им необходим контакт с внеклеточным матриксом соединительной ткани, и этот контакт реализуется путем взаимодействия белков внеклеточного матрикса с интегринами [2, 4]. Как показано на схеме, интегрины участвуют в активации белков Ras — центральных внутриклеточных протеинкиназ сигнальных путей МАРК (МАРК — mitogen activated protein kinase — киназы, активируемые митогенными факторами и приводящие к митозу клеток). По последним данным, гормоны щитовидной железы, в частности, тетрайодтиронин, служит лигандом интегриновых ICAM-белков (белков межклеточной адгезии), тем самым способствуя пролиферации клеток.
Это во многом определяет важную роль гормонов щитовидной железы в развитии нервной системы не только в зародышевом периоде, но и у детей, и объясняет задержку психомоторного развития при врожденном гипотиреозе — первичной недостаточности щитовидной железы. С другой стороны, это объясняет повышенный риск возникновения и прогрессирования злокачественной опухоли на фоне гипертиреоза [11].
Бывает так, что в генах, кодирующих участников этой передачи ростового сигнала, происходит мутация — стойкое и необратимое изменение структуры того или иного гена. Это происходит при воздействии самых разных факторов: физических (радиация в виде ионизирующего излучения) или химических (нитриты, канцерогены в примесях сигарет, различные фенолы). Отдельное значение имеют биологические факторы канцерогенеза. К таковым на сегодняшний день относят вирус Эпштейна — Барр, онкогенные типы вируса папилломы человека (16, 18, 31 и целое множество других), бактерию Helicobacter pylori. При сочетании этих канцерогенных факторов, скажем, с дефектом системы репарации (системы «починки» имеющихся дефектов) или системы апоптоза (генетически детерминированной гибели клетки) или с каким-то другим вариантом молекулярно-генетических нарушений возникает очень опасная ситуация — функциональная аномалия клетки, выражающаяся в ее автономии. Теперь такая клетка живет и растет только по своим законам. Происходит гиперпродукция и/или чрезмерная спонтанная активность определенного участника митогенных внутриклеточных каскадов, обусловливающих полную клеточную независимость от каких-либо внешних сигналов и отрицательной обратной связи.
Протоонкогены
В нормальных условиях протоонкогены обеспечивают выживание клетки, когда это необходимо в рамках гистогенеза и строго при воздействии митогенных сигналов извне.
Чтобы продемонстрировать, как все это происходит, я предлагаю углубиться в дебри молекулярной онкологии.
Как это работает? На рис. 22 можно заметить сигнальный каскад, начинающийся от RTK (ТКР). RTK — это тирозинкиназный рецептор (ТКР), т. е. рецептор, обладающий тирозинкиназной активностью. Но приобретает он ее только после своей активации.
При воздействии лиганда (факторы роста) на ТКР происходит димеризация (удвоение) двух тирозинкиназных рецепторов. В активированном состоянии ТКР фосфорилирует по остаткам тирозина нижележащие участники сигнального каскада, в частности, белки SOS и GRB2. Далее указанные белки фосфорилируют нижележащие белки — RAS → RAF→ MEK→ ERK (в соответствии с рис. 22).
Последовательная активация данных белков заканчивается фосфорилированием транскрипционных факторов, запускающих экспрессию генов, необходимых для пролиферации.
В опухолевых клетках происходит все то же самое, что и в обычных, с одним отличием: если нормальным клеткам для запуска пролиферации нужен сигнал извне, то опухолевая клетка сама себе хозяин.
Так, при мутации гена EGFR (наиболее известный ТКР) активность рецептора сохраняется постоянно, даже в отсутствии лиганда, что приводит к постоянной активации сигнального каскада, описанного выше. Более того, поломки могут происходить на любом участке каскада: мутации могут возникать в гене, кодирующем белки RAS, RAF, MEK и множество других, и результат будет тот же самый: неконтролируемая пролиферация клетки. Мутации гена EGFR возникают в некоторых случаях аденокарциномы легкого, плоскоклеточного рака кожи, мутации RAS — при раке поджелудочной железы, некоторых случаях рака толстой кишки, некоторых случаях немелкоклеточного рака легкого, мутации RAF — при той же аденокарциноме легкого, меланоме, в некоторых случаях рака толстой кишки. Список можно продолжать бесконечно, потому что подобных сигнальных путей в клетках очень много. В случае каждой конкретной опухоли «ломается» один или несколько сигнальных путей. Все это создает почву для создания таргетной терапии. Таргетные препараты нацелены на инактивацию конкретного белка, опосредующего онкогенез, тем самым достигается остановка опухолевого роста, а в ряде случаев даже уменьшение имеющихся опухолевых очагов.
К наиболее известным протоонкогенам относят [2, 4, 6]:
- тирозинкиназные рецепторы к факторам роста (EGFR, FGFR, PDGFRA, IGF, c-KIT, VEGFR A-C и прочие): при мутации в данных генах происходит постоянная активация нижележащих сигнальных путей, приводящих к пролиферации клеток [12]. Их мутации лежат в основе патогенеза различных опухолевых заболеваний.
- мутации в «нижележащих точках» сигнальных каскадов, приводящие к гиперэкспрессии генов внутриклеточных участников трансдукции сигнала (на рис. 27 это белки RAS, BRAF, МЕК, а также множество других, которые просто не поместятся в эту статью).
- антиапоптотические факторы, например, Bcl- 2 [12].
Зачем это знать? Многие из вышеперечисленных белков являются мишенями для препаратов таргетной терапии (целенаправленная терапия). Существующие препараты сильно поменяли тактику лекарственного лечения ряда опухолевых заболеваний.
В качестве наиболее распространенных примеров: аденокарцинома легкого при наличии мутаций гена EGFR поддается лечению ингибиторами EGFR (осимертиниб, гефитиниб, афатиниб — в зависимости от вида мутации). Меланома при мутации RAF (в частности, BRAFV600E) поддается лечению BRAF-ингибиторами (вемурафениб, дабрафениб). Более подробно мы расскажем об этом в другой раз.
Онкосупрессоры
Иногда происходит так, что мутация задевает гены, кодирующие белки-онкосупрессоры. Это белки, способные заблокировать клеточный цикл в случае наличия неисправимых повреждений ДНК и включить программу апоптоза.
К наиболее известным белкам-онкосупрессорам относят p53, pRb, PTEN. Их функции различны. Так, р53 запускает программу апоптоза путем активации каспаз 3, 8, 9 и других молекул-участников апоптоза. РRb, или белок ретинобластомы, останавливает движение клеточного цикла в фазе G1, не давая клетке пролиферировать (рис. 23).
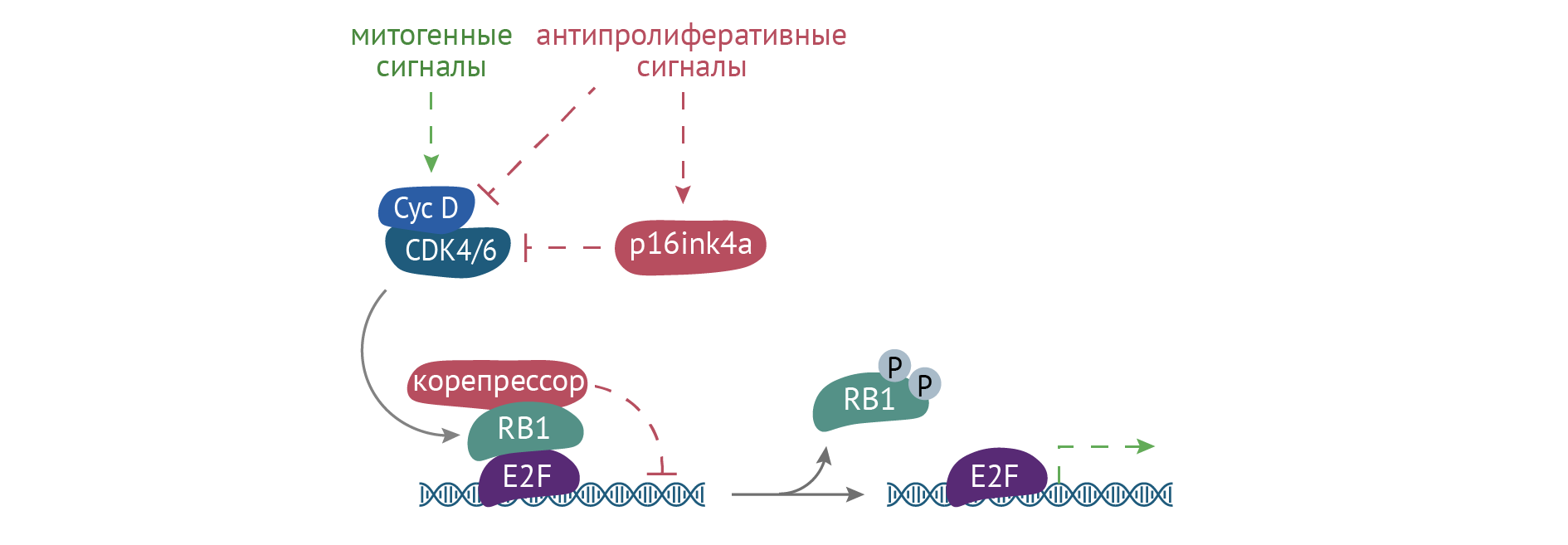
Рисунок 23 | Переход из G1 в S фазу клеточного цикла
Белок ретинобластомы (RB1) выступает контрольной точкой между этими двумя фазами. При фосфорилировании pRB1 распадается комплекс RB1-E2F, который тормозил прохождение клеткой клеточного цикла. Тем самым E2F в отсутствие угнетающего влияния RB1 запускает экспрессию генов, необходимых для репликации ДНК.
Происходит это за счет инактивации соответствующих факторов репликации, в частности, EEF2 (эукариотический фактор элонгации 2). PTEN (рис. 28) является инактиватором различных участников МАРК-путей. Нарушение работы онкосупрессоров в результате мутации их гена — важный фактор формирования большинства опухолей у человека [2, 8, 12], а также предиктор неблагоприятного исхода онкологического заболевания.
В результате мутаций в протоонкогенах и генах-онкосупрессорах мы получаем клетки, неадекватно реагирующие на ростовые сигналы. Такие клетки способны к автономному росту, инвазии в окружающие ткани, а затем — к метастазированию в отдаленные уголки организма [2]. Все это составляет суть опухолевого процесса.
Биохимические аспекты цитостатической терапии, направленной на угнетение метаболизма ДНК
Химиотерапия злокачественных новообразований различной природы на примере ингибиторов топоизомераз
На сегодняшний день достигнуты значительные успехи в лечении злокачественных новообразований. Разработаны схемы и режимы цитостатической химиотерапии для опухолей различной локализации и морфологической природы. Несмотря на различие этих схем, принцип действия химиотерапевтических цитостатических препаратов в глобальном и общем смысле является универсальным. Он заключается в остановке клеточного цикла и угнетении пролиферации опухолевых клеток. Каждая группа препаратов делает это по-своему. Главным недостатком цитостатической химиотерапии является низкая селективность препаратов: они угнетают все пролиферирующие клетки организма.
Ингибиторы топоизомераз. В опухолевых клетках в силу высокой пролиферативной активности прослеживается высокая активность топоизомераз (преимущественно I и II типов). Теканы (топотекан, иринотекан) ингибируют топоизомеразу I типа. Антрациклины (доксорубицин, идарубицин эпирубицин), эпиподофиллотоксины (производные подофиллотоксина, такие как этопозид, тенопозид) являются ингибиторами топоизомераз II типа. Поскольку топоизомеразы поддерживают пространственную ориентацию нитей ДНК при репликации, их ингибирование является фатальным для опухолевой клетки. Указанные выше группы препаратов нашли широкое применение в онкологии. Так, антрациклины входят в схемы лечения солидных опухолей (в частности, рака легкого, желудка, рака молочной железы, мочевого пузыря, сарком мягких тканей), онкогематологических заболеваний [3, 7].
К сожалению, данные типы топоизомераз присутствуют и в здоровых тканях организма. Из-за этого развиваются побочные эффекты, часто специфичные для некоторых ингибиторов топоизомераз. Так, в последние годы появляется все больше сообщений о развитии антрациклиновой кардиотоксичности.
Антрациклин-индуцированная кардиотоксичность — феномен, характеризующийся постоянно прогрессирующим снижением фракции выброса левого желудочка миокарда с последующим развитием хронической сердечной недостаточности. Развитие данного осложнения связано с тем, что механизм противоопухолевого действия антрациклинов включает не только ингибирование топоизомеразы опухолевых клеток, но и продукцию в них активных радикалов кислорода, разрушительно влияющих на ДНК.
Опасность антрациклиновой кардиотоксичности заключается и в том, что зачастую очень сложно отследить развитие сердечной недостаточности в силу ее поздней клинической манифестации. Между тем Европейское общество кардиологов (ESC), а также Европейское общество медицинской онкологии (ESMO) предложили рекомендации для ведения пациентов, получающих кардиотоксичную химиотерапию. Более подробно об этом можно узнать здесь.
Химиотерапия аутоиммунных заболеваний на примере антиметаболитов
Механизмы развития аутоиммунных заболеваний сложны, но достаточно часто они включают в себя пролиферацию аутореактивных (опасных в отношении тканей собственного организма) лимфоцитов. Это обусловлено угнетением негативной селекции аутореактивных лимфоцитов, избеганием ими апоптоза. Лимфоциты относятся к группе быстро пролиферирующих клеток, поэтому в лечении множества аутоиммунных (иммунопатологических) заболеваний применяются цитостатики. Наиболее известными являются антиметаболиты фолиевой кислоты, в частности, метотрексат [10].
Метотрексат является ингибитором дигидрофолатредуктазы и тимидилатсинтетазы (рис. 24)
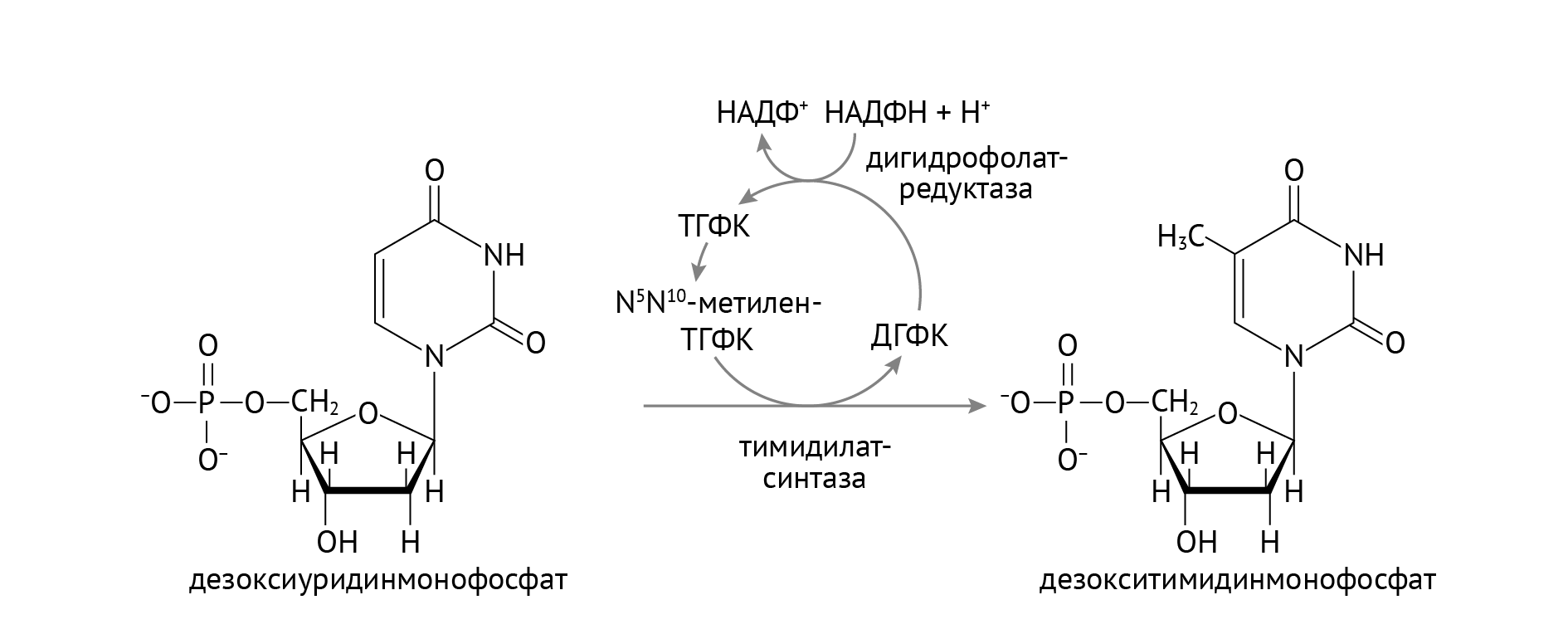
Рисунок 24 | Образование дТМФ. Тетрагидрофолатредуктаза как мишень для метотрексата.
Конкурентное ингибирование данных ферментов приводит к снижению синтеза одного из субстратов, необходимых для репликации ДНК — дезокситимидинтрифосфата. Механизм действия обусловливает применение метотрексата в лечении как опухолевых заболеваний (преимущественно гемобластозов), так и аутоиммунных, например, ревматоидного артрита, болезни Крона и неспецифического язвенного колита (один из препаратов резерва), а также диффузных заболеваний соединительной ткани. Хотя сейчас появились более безопасные и эффективные препараты, метотрексат и по сей день не потерял своего значения. Особое место он занимает в лечении ревматоидного артрита — аутоиммунного поражения суставов и периартикулярных тканей, которое без лечения заканчивается контрактурой суставов, ограничением подвижности, инвалидностью, амилоидозом внутренних органов и множеством других проблем.
Подведем итоги
Синтез ДНК, процесс считывания и реализации генетической информации таит в себе множество вопросов. Однако именно в этих сложных процессах можно найти ключ к лечению многих заболеваний. Для меня это был самый интересный раздел за все годы обучения в университете, потому что это фундаментальный вопрос, в котором можно копаться бесконечно. Коснувшись процессов репликации ДНК и синтеза белка, мы с тобой попутно рассмотрели вопросы антибактериальной терапии, аспекты возникновения опухолей и их лечения, а также терапию аутоиммунных заболеваний, и это я вовремя остановился. Я осветил лишь те моменты, которые казались мне наиболее интересными и важными, но, если у меня получилось заинтересовать тебя и побудить разбираться самостоятельно, значит, я писал все это не напрасно.
В молекулярной биологии можно найти объяснение очень многим вещам, но у меня пока получается находить только все новые и новые вопросы.
Глава 3 | Катаболизм и биоэнергетика
Глава 4 | Катаболизм липидов
Источники
- Наглядная биохимия / Я. Кольман, К.-Г. Рём ; пер. с англ. Т. П. Мосоловой. — 5-е изд., перераб. и доп. — М. : Лаборатория знаний, 2018.
- Патологическая биохимия /Таганович А.Д., Олецкий Э.И., Котович И.Л./Под общей редакцией Тагановича А.Д. – М.: БИНОМ, 2015 г..
- Нельсон Д., Кокс М. Основы биохимии Ленинджера: в 3 т. Т. 3. – М.: Лаборатория знаний, 2015 г.
- Тараховский Ю.С., Ким Ю.А., Абдрасилов Б.С., Музафаров Е.Н. Флавоноиды: биохимия, биофизика, медицина, 2013 г.
- «Регуляция активности протеасом в клетке с помощью специфического выключения гена, кодирующего одну из субъединиц протеасомы метода РНК-интерференции». Монахов М.В, Институт Молекулярной биологии им. В.А. Энгельгардта РАН, 2004 г.
- Тимин О.А. Основы биологической химии, 2018 г.
- Cristiana Sessa, Luca Guanni et al. ESMO HANDBOOK OF CLINICAL PHARMACOLOGY OF ANTI-CANCER AGENTS, 2012.
- Портал «Биохимия для студента», раздел «Матричные биосинтезы». URL: https://biokhimija.ru/lekcii-po-biohimii.html.
- В. М. Виноградов, Е. Б. Каткова. Фармакология с рецептурой, 2016 г.
- Федеральные клинические рекомендации «Ревматоидный артрит», 2017 г.
- Глушаков Р.И., Власьева О.В., Соболев И.В., Прошин С.Н., Тапильская Н.И. Тиреоидный статус как прогностический маркер в онкологии. Злокачественные опухоли, 2015.
- C. Зилбернагль, Ф. Ланг. Клиническая патофизиология, 2016 г.
- Serrato-Capuchina A, Matute DR. The Role of Transposable Elements in Speciation. Genes (Basel), 2018. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5977194/
- Ou HD, Phan S, Deerinck TJ, Thor A, Ellisman MH, O'Shea CC. ChromEMT: Visualizing 3D chromatin structure and compaction in interphase and mitotic cells. Science. 2017 Jul 28. https://doi.org/10.1126/science.aag0025
- Mismatch repair deficiency/microsatellite instability-high as a predictor for anti-PD-1/PD-L1 immunotherapy efficacy Pengfei Zhao, Li Li, Xiaoyue Jiang, and Qin Li J Hematol Oncol. 2019; 12: 54. Published online 2019 May 31. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6544911
- The ENCODE Project Consortium., Overall coordination (data analysis coordination)., Dunham, I. et al. An integrated encyclopedia of DNA elements in the human genome. Nature 489, 57–74 (2012). https://doi.org/10.1038/nature11247
- Sendzik J, Lode H, Stahlmann R. Quinolone-induced arthropathy: an update focusing on new mechanistic and clinical data. Int J Antimicrob Agents. 2009 Mar;33(3):194-200. https://doi.org/10.1016/j.ijantimicag.2008.08.004
- van der Linden PD, Sturkenboom MCJM, Herings RMC, Leufkens HMG, Rowlands S, Stricker BHC. Increased Risk of Achilles Tendon Rupture With Quinolone Antibacterial Use, Especially in Elderly Patients Taking Oral Corticosteroids. Arch Intern Med. 2003;163(15):1801–1807. https://doi.org/10.1001/archinte.163.15.1801
- Guan MX, Fischel-Ghodsian N, Attardi G. A biochemical basis for the inherited susceptibility to aminoglycoside ototoxicity. Hum Mol Genet. 2000 Jul 22;9(12):1787-93. https://doi.org/10.1093/hmg/9.12.1787
- Reilly MT, Faulkner GJ, Dubnau J, Ponomarev I, Gage FH. The role of transposable elements in health and diseases of the central nervous system. J Neurosci. 2013;33(45):17577-17586. https://dx.doi.org/10.1523%2FJNEUROSCI.3369-13.2013
